Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
| Code | CSB-EP019898HU |
| Abbreviation | Recombinant Human RNF8 protein |
| MSDS | |
| Size | $256 |
| Order now | |
| Image |
|
| Have Questions? | Leave a Message or Start an on-line Chat |
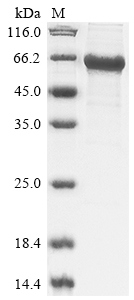
The process for recombinant Human RNF8 protein synthesis in e.coli cells includes recombining the DNA fragment that encodes the Human RNF8 protein (1-485aa) into expression vector, transforming the recombinant vector into e.coli cells, screening positive cells, cultivating and inducing cells, conducting cell lysis, and performing expression analysis. The resulting recombinant Human RNF8 protein is purified from the cell lysate using affinity purification, with its purity exceeding 85% as determined by SDS-PAGE.
There are currently no reviews for this product.

