Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
| Code | CSB-MP773799HU |
| Abbreviation | Recombinant Human BTLA protein, partial (Active) |
| MSDS | |
| Size | $138 |
| Order now | |
| Image |
|
| Have Questions? | Leave a Message or Start an on-line Chat |

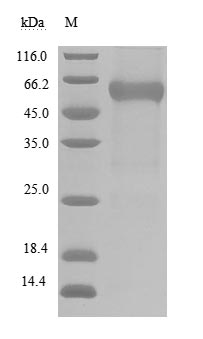
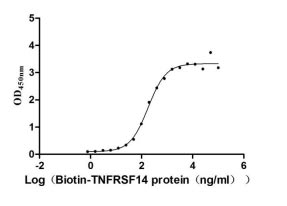
BTLA (CD272) functions as a coinhibitory receptor in the immune checkpoint network, engaging HVEM (TNFRSF14) to dampen T- and B-cell activation—making it a compelling target in immuno-oncology drug discovery. This recombinant human BTLA encompasses the extracellular ligand-binding domain (aa 31–150) with a C-terminal hFc1-Myc tag, and mammalian cell expression preserves the native folding and post-translational modifications critical for faithful receptor-ligand interaction. Functional ELISA confirms robust binding to biotinylated human TNFRSF14 with an EC50 of 137.8–233.4 ng/ml, providing validated support for ligand-binding assays, competitive inhibition studies, blocking antibody screening, and therapeutic antibody epitope mapping. Purity exceeding 90% by SDS-PAGE and endotoxin levels below 1.0 EU/μg satisfy the quality thresholds commonly required for SPR- or BLI-based affinity characterization, small-molecule inhibitor screening, and use as a positive control in checkpoint receptor binding assays.
There are currently no reviews for this product.

