Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
Exosomes were originally regarded as cellular metabolic waste. With the discovery in 1996 that B lymphocyte-derived exosomes were reported to present antigens [1] and the exchange of genetic material between cells via exosomal RNA in 2007 [2], people became aware of the importance of exosomes. These findings have also spurred scientists to explore and study exosomes. Exosome research has become a research hotspot since 2007 [3][4].
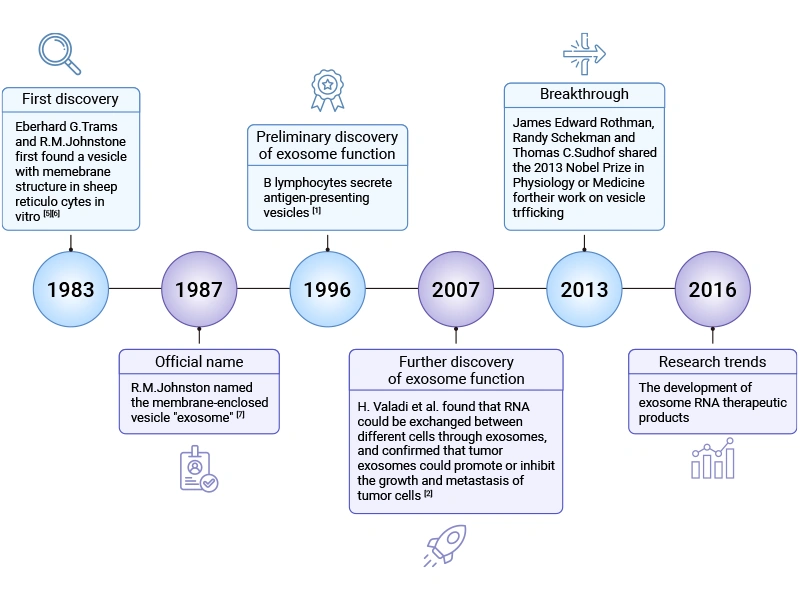
Figure 1. The development history of exosome research
This article mainly describes the definition of exosomes, the importance of exosome isolation, methods for exosome separation, identification of exosome quality and purity, as well as troubleshooting and best practices in exosome isolation.
Table of Contents
2. Why Do People Isolate Exosomes?
3. Core Methods for Isolating Exosomes
4. Assessing Exosome Quality and Purity
Exosomes, a specific subtype of extracellular vesicles (EVs), are nano-sized (typically 30-150 nm in diameter) lipid-bilayer particles naturally secreted by most eukaryotic cells [8]. They are released through the endosomal pathway and carry proteins, lipids, and nucleic acids from their cell of origin. Exosomes play essential roles in intercellular communication by transferring their cargo between cells, influencing physiological and pathological processes, especially in neurodevelopment, immune modulation, and disease progression.
![Typical representative composition of exosomal cargo [14]](https://www.cusabio.com/manage/upload/202604/Exosomal-cargoes.webp)
Figure 2. Typical representative composition of exosomal cargo [14]
Exosome biogenesis initiates with the inward budding of the plasma membrane, which internalizes soluble extracellular molecules and membrane-associated proteins to form early sorting endosomes (ESEs). During maturation, the limiting membrane of these early endosomes undergoes invagination, leading to the formation of multivesicular bodies (MVBs). Within MVBs, specific biomolecules and particles are selectively sorted and encapsulated into intraluminal vesicles (ILVs), a critical step that determines the cargo composition and functional properties of future exosomes. The MVBs are finally trafficked to either fuse with lysosomes for degradation or with the plasma membrane to release the ILVs into the extracellular space as exosomes [8].
Exosomes' immense potential across several transformative fields has driven exosome isolation to become a cornerstone of modern biomedical research. The primary motivations for isolating exosomes stem from their unique biological roles, their composition reflective of parental cells, and their accessibility in bodily fluids, positioning them as powerful tools for diagnosis, therapy, and fundamental biological discovery.
The isolation of high-purity, intact exosomes from complex biological matrices (e.g., cell culture supernatant, serum, plasma, cerebrospinal fluid) is a prerequisite for downstream basic research, biomarker discovery, and therapeutic development.
One of the most compelling reasons to extract exosomes is their utility in liquid biopsy. Exosomes are abundant in easily accessible biofluids like blood (serum/plasma), urine, and cerebrospinal fluid [9-11]. Their cargo, such as proteins, lipids, DNA, and various RNA types (mRNA, microRNA, lncRNA), largely reflects the molecular state of their cell of origin, including diseased cells like cancer cells [10,12,13]. By isolating exosomes from patient samples, researchers can analyze this cargo for specific signatures that indicate the presence, type, stage, or prognosis of a disease [14,15].
For instance, specific exosomal microRNAs have been identified as promising biomarkers for discriminating between patient groups, such as those with metastatic prostate cancer, treated cancer, and healthy controls [10]. Isolating exosomes enables the profiling of these miRNAs with high sensitivity and specificity, offering a minimally invasive alternative to tissue biopsies [16,17].
Exosomes are natural intercellular communicators, involved in signal transduction and maintaining homeostasis [9,13]. This inherent biological function makes them attractive as novel therapeutic agents and drug delivery vehicles. Isolated exosomes can be engineered to carry therapeutic cargo (e.g., drugs, nucleic acids) and targeted to specific tissues or cells [16].
Research explores the use of exosomes for regenerative medicine, such as applying mesenchymal stem cell-derived exosomes to promote tissue repair, leveraging effects similar to those of the source cells without the risks of cell transplantation [18]. Furthermore, isolating exosomes following specific stimuli (such as shock wave therapy) allows scientists to study and potentially harness their inherent regenerative and angiogenic potential for treating conditions ranging from tendinopathies to ischemic heart disease [19]. Effective isolation is the critical first step in developing such exosome-based therapeutics [14,16].
Exosomes are key mediators of intercellular communication, playing essential roles in both normal physiology and the pathogenesis of diseases [13-15]. By extracting and analyzing exosomes from patients, researchers can decipher the messages that cells send to one another in contexts ranging from immune responses and tissue crosstalk to cancer metastasis [15,20].
For example, isolating exosomes from cell culture supernatants enables detailed characterization of their proteome, providing insights into the molecular reprogramming they induce in recipient cells [20]. Similarly, comparing exosomes from different biological states (e.g., from vaccinated versus tumor-bearing subjects) can reveal crucial differences in RNA and protein content, shedding light on underlying mechanisms of protection or disease progression [21].
Extraction is a prerequisite for any subsequent analytical or clinical procedure. The choice of isolation method directly impacts the yield, purity, and integrity of the obtained exosomes, which in turn affects the reliability of downstream analyses like RNA sequencing, proteomics, functional assays, and biomarker validation [17,20,22,23].
Whether the goal is to perform sensitive miRNA profiling, identify vesicle-specific proteins by mass spectrometry [20], or test exosome function in cell proliferation and migration assays, a robust and reproducible isolation protocol is essential. The drive towards standardization in exosome extraction methods is fundamentally linked to the need for consistent, high-quality samples that can be reliably used in clinical settings [23,24].
Exosomes must be separated from non-exosomal components, such as microvesicles, apoptotic bodies, and other biomolecules, in sufficient quantities, with high purity, and of appropriate size to support fundamental research. The best strategy for separating exosomes should be chosen based on the specific type of bodily fluid or tissue used as the source. However, no single method is ideal for all applications, so most laboratories match the technique to their main question, available equipment, and tolerance for impurities.
Traditional methods for exosome isolation have been developed and refined over time, each with distinct principles, advantages, and inherent limitations that influence the yield, purity, and integrity of the isolated vesicles.
Table 1 summarizes the common exosome isolation methods, including their separation principles, characteristics, common sample types, time required, advantages, and disadvantages [14].
| Method | Principle | Advantage | Disadvantage | Recovery | Purity | Time | Scalability | Sample volume | Principle Schematic | |
|---|---|---|---|---|---|---|---|---|---|---|
| Ultracentrifugation | Differential ultracentrifugation (DUC) | Different particles have different sedimentation coefficients | • Suitable for large-volume samples • No other markers will be introduced • Low cost |
• High equipment • Labor-intensive • Potential destruction of exosomes • Exosome aggregation |
High | Low | ≈4 h | Low | Large |
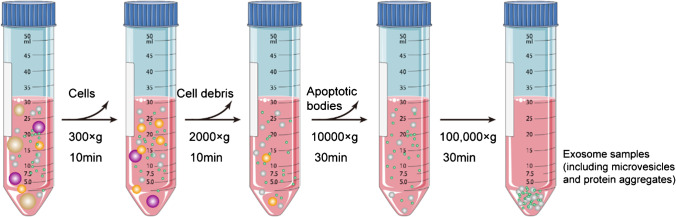
Figure 3. Schematic of DGUC–based exosome isolation [14] |
| Density gradient ultracentrifugation (DGUC) | Components with imparity of size and density possess various sediment speed | • Higher purity vs. DUC • Separate exosomal subpopulation |
• Tedious operation • Limited treatment capacity • Low throughput |
Low | High | 10–18 h | Medium | Medium |
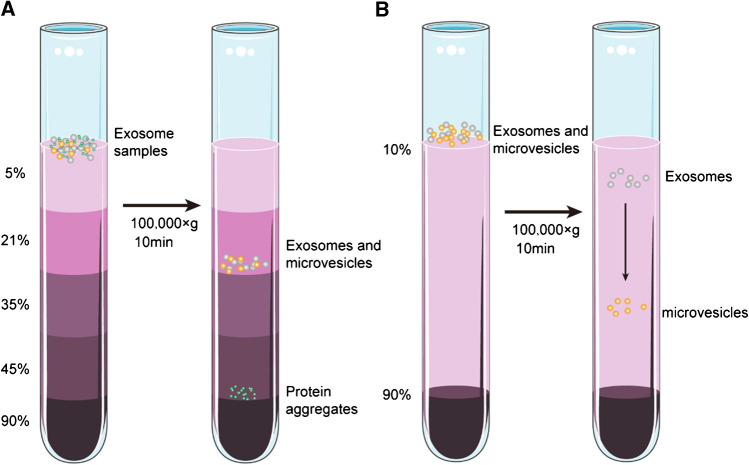
Figure 4. Schematic of DGUC–based exosome isolation [14] |
|
| Ultrafiltration (UF) | Particles with various size and molecular weight (Use a membrane with a specified pore diameter or MWCO) | • Low cost • Time efficient • Simple |
• Rupture of exosome • Membrane blockage |
Medium | Medium | ≈2–4 h | High | Medium |
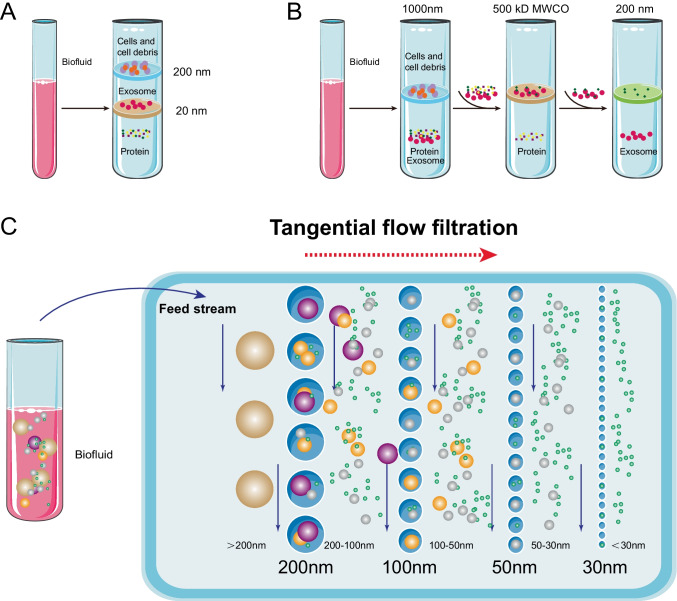
Figure 5. Schematic of DGUC–based exosome isolation [14] |
|
| Size exclusion chromatography (SEC) | Separates exosomes based on hydrodynamic radii | • Preserve the integrity and natural biological
activity • No additional preprocessing |
• Potential contamination • High equipment cost |
High | High | 15 min | High | Small |
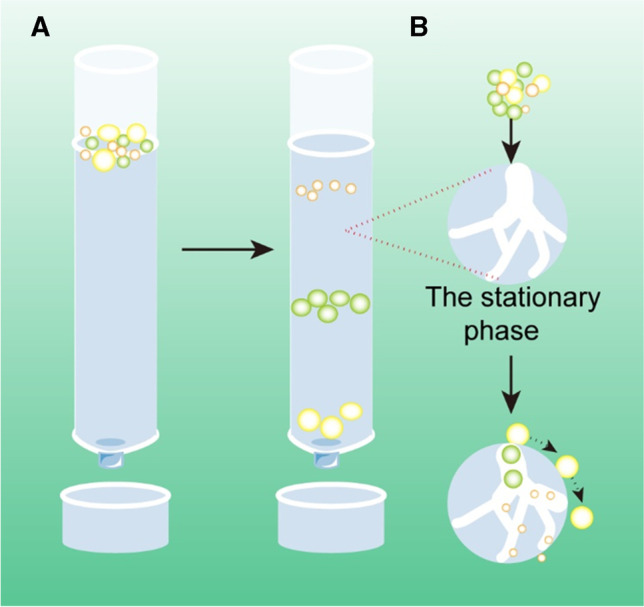
Figure 6. Schematic of DGUC–based exosome isolation [14] |
|
| Immunoaffinity capture | Based on interaction between antibodies and specific membrane proteins of exosomes | • High specificity • Simple |
• Destroy the integrity of the exosome • Expensive • Nonspecific binding |
Low | High | 2–6 h | High | Small |
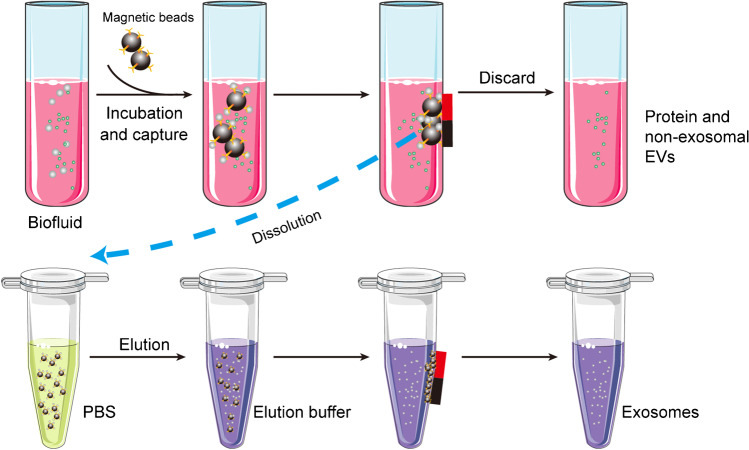
Figure 7. Schematic of DGUC–based exosome isolation [14] |
|
| Polymer precipitation | Polymer can adhere and precipitate exosomes | • Suitable for all types of samples • Simple and rapid • No deformation of the exosomes |
• Lead to the wrong quantification • Additional steps for higher purity |
High | Medium | 0.5–12 h | Medium | Small |
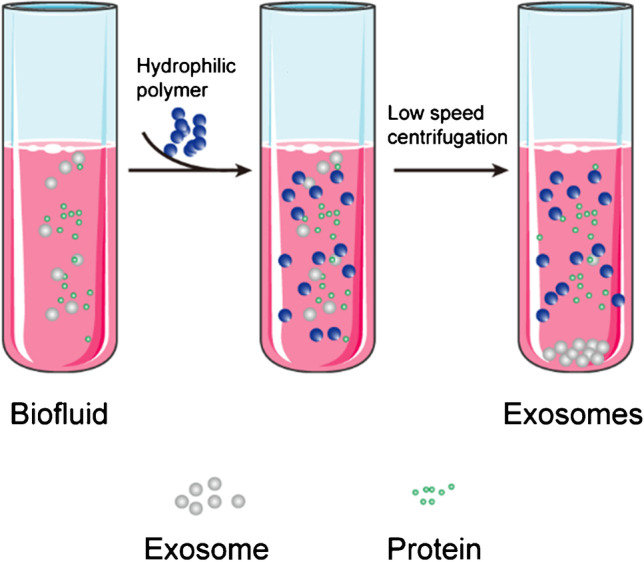
Figure 8. Schematic of DGUC–based exosome isolation [14] |
|
The isolation of exosomes, crucial for downstream analyses in diagnostics and biomarker discovery, presents significant challenges regarding purity, yield, and integrity. Microfluidic technologies have emerged as powerful platforms to address these challenges, offering advantages such as low sample/reagent consumption, rapid processing, and potential for integration and automation. These techniques can be broadly categorized into passive (label-free) and active (or affinity-based) methods, each with distinct operational principles and performance characteristics.
Table 2: Comparison of Microfluidic-Based Exosome Isolation Methods
| Isolation Method | Classification | Working Principle | Advantages | Limitations |
|---|---|---|---|---|
| General Microfluidic Technique (based on fluid properties) | Modular platform compatible with diverse exosomal isolation strategies | Utilizes microchannels that can be flexibly connected according to specific application requirements, enabling integration with various exosomal isolation methods for high-throughput processing [27] | Low consumption of sample volume and reagents; Short isolation time [28]; Synergistically improves exosome yield and purity when combined with other isolation methods |
Risk of channel blockage by analytical samples; Low exosome output due to limited sample loading capacity; Requires advanced equipment, limiting large-scale use |
| Size-based Microfluidic Isolation Technique | Passive, label-free isolation method | Separates exosomes based on size differences, mainly via size-sieving structures (e.g., membranes with tunable pore sizes, nanowire-based systems) to isolate exosomes within a specific size range [29, 30, 37, 38] | High-throughput potential; ExoTIC: Simple, high recovery & purity from small samples (10–100 µL), suitable for clinical testing [29, 37]; Nanowire/DLD Arrays: Label-free, specific size cut-off possible [30, 32, 38] |
Purity can be compromised (similar to SEC); Potential issues with exosome saturation and low recovery in some designs [32]; Risk of device blockage |
| Deterministic Lateral Displacement (DLD) Technique | Passive, label-free isolation method | Utilizes a cylindrical gradient array with a defined critical particle size; particles larger than the critical size have their flow path altered, while smaller particles (below the critical size) maintain their original flow path, achieving size-based separation [39] | Label-free and easy to operate for on-chip exosome separation; Can separate exosomes from other EVs [31]; enables chip-based exosome sorting and quantification [32] |
Limited by exosomal saturation and low recovery rate [32]; Risk of channel clogging, leading to low isolation purity; |
| Immune Microfluidic Technique | Label-based isolation method | Isolates exosomes via specific binding between exosome surface markers and antibodies immobilized on the surface of microfluidic chips | High isolation efficiency; Fast processing speed; Simple operation; Low sample volume requirement; |
The integrity and purity of isolated exosomes need to be improved; Non-specific binding is a core bottleneck, as blocking and washing steps are difficult to perform in microfluidic devices; Acidic elution for antibody-bound exosome release may contaminate samples and damage exosomes |
| Dielectrophoretic (DEP) Technique | Active, label-free isolation method | In a non-uniform electric field, particles become polarized and are subjected to a dielectric force correlated with their size (inversely proportional to particle radius) and electrical properties; size-dependent exosome isolation is achieved by capturing particles via a gradient of the square electric field [33, 40] | Label-free, non-contact, rapid, and high-throughput; Enables simultaneous capture and separation of exosomes; Can be integrated with immunoaffinity detection to achieve a low limit of detection and a wide detection range [34]; |
Joule heating and electrothermal heating effects require optimized electrode design to avoid impacts on isolation performance [41]; Low separation resolution and purity; Few relevant studies on exosome isolation; The functional integrity of isolated exosomes remains to be verified, requiring further in vitro studies and downstream analyses (e.g., RNA sequencing, proteomics) |
| Acoustofluidic Isolation | Active, label-free isolation method | Uses acoustic waves (e.g., surface acoustic waves) to exert forces on particles; particles with different mechanical properties (size, density) are separated in the sound field | Enables automated, direct isolation of exosomes from undiluted whole-blood samples; High isolation purity (up to 98.4% from EV mixtures); Significantly higher yield of exosomal small RNA compared to differential ultracentrifugation (DUC) [35, 36] |
Mainly applied to isolation of larger objects (e.g., cells, bacteria); precise manipulation of particles below 100 nm (exosomes) remains technically challenging, with few breakthroughs in exosome-specific isolation |
| Asymmetric Flow Field-Flow Fractionation (AF4) | Passive isolation method | A size-based separation technique with programmable cross-flow intensity (distinct from size-exclusion chromatography, SEC), which can be optimized for exosome isolation to separate EV subsets based on size differences [42] | Enables high-reproducibility and high-purity exosome separation [43, 44]; Preserves the integrity of exosomes; Programmable cross-flow allows optimization for different exosome sizes [42]; Powerful for subclassification of EVs [25,26] |
Low resolution |
Commercial exosome isolation kits have emerged as scalable, user-friendly alternatives for exosome extraction, with diverse technical designs tailored to different research priorities such as yield, purity, throughput, and sample compatibility.
Table 3: Comparison of Commercial Exosome Isolation Kits
| Kit Category | Core Working Principle | Key Advantages | Critical Limitations |
|---|---|---|---|
| Polymer Precipitation-Based Kits | Hydrophilic polymers (primarily PEG with molecular weights of 6000–8000 Da) compete for water molecules in the sample to reduce exosome solubility, driving exosome aggregation and precipitation via low-speed centrifugation (1,000–3,000 × g) | Ultra-high exosome yield (80–300× higher than ultracentrifugation) [46]; Simple workflow, no specialized equipment, <2h processing time, with compatibility for 96-well plate formats for large cohort studies [47]; Robust performance across sample types and exosomal RNA/miRNA recovery; Low per-sample cost compared to chromatography or affinity-based kits |
Severe non-specific co-precipitation of abundant soluble proteins (e.g., albumin), lipoproteins (LDL, HDL), and protein aggregates [48]; Incompatibility with high-purity proteomics applications: PEG contamination and co-precipitated proteins interfere with mass spectrometry-based proteomic analysis, distorting protein quantification and reducing the detection of low-abundance exosomal markers [46]; Impaired functional assay performance: PEG-precipitated exosome preparations exhibit reduced stability in biological assays, as PEG can induce cytotoxicity in cell cultures and co-isolated contaminants cause non-specific bioactivity [46]; Cannot separate exosomes from similarly sized non-exosomal EVs/protein complexes, resulting in broad particle size distributions and reduced specificity [46] |
| Size Exclusion Chromatography (SEC)-Based Kits | Porous gel filtration resin columns separate exosomes from contaminants based on hydrodynamic size; exosomes (30–150 nm) elute in the void volume first, while small soluble proteins/lipoproteins are retained in resin pores | Superior exosome purity with minimal soluble protein contamination [48]; Mild, physiological separation conditions preserve exosome morphology, membrane integrity and biological activity; High reproducibility across samples and laboratories; Scalable formats compatible with sample volumes from 100 μL to 50 mL |
Moderate exosome yield compared to precipitation kits [48]; Cannot fully separate exosomes from size-overlapping VLDL/LDL lipoproteins in plasma samples; Final exosome preparations are diluted, requiring additional concentration steps; Higher per-sample cost than precipitation kits, limited high-throughput compatibility |
| Immunoaffinity Capture-Based Kits | Antibodies targeting exosome surface markers (canonical tetraspanins CD9/CD63/CD81 or cell-type-specific markers) are immobilized on solid supports (magnetic beads/resins) to capture target exosomes via highly specific antigen-antibody interactions | Exceptional specificity and purity, minimal co-isolation of non-exosomal contaminants; Enables isolation of cell-type/tissue-specific exosome subpopulations from complex biofluids; Mild conditions preserve exosome structural and functional integrity; Streamlined workflow compatible with direct downstream analysis without elution |
Low total exosome yield, limited to vesicles expressing the target marker(s); High per-sample cost from antibody-coated consumables, not suitable for large cohort studies; Harsh elution may damage exosome proteins, while mild elution leads to incomplete recovery; Marker expression bias across cell types/disease states may distort comparative results |
| Ultrafiltration (UF)-Based Membrane Affinity Kits | Centrifugal spin columns with 100 kDa molecular weight cut-off membranes (MWCO) retain and concentrate exosomes, while low-molecular-weight proteins/metabolites pass through the membrane; advanced versions integrate membrane affinity chemistry to enhance exosome binding | Rapid concentration and isolation: Simultaneous isolation and concentration of exosomes in <1 hour, no specialized equipment [47]; High recovery of intact exosomes: Mild centrifugal conditions avoid exosome aggregation and membrane damage; Compatible with diverse downstream omics and functional assays; Flexible formats accommodate sample volumes from 250 μL to 50 mL |
Membrane fouling causes significant non-specific protein contamination; Risk of exosome loss and damage via membrane pore penetration, or deformation/rupture from prolonged centrifugation [47]; Cannot separate exosomes from similarly sized EVs, lipoproteins or protein aggregates; Batch-to-batch variability in membrane performance |
| Phosphatidylserine (PS) Affinity-Based Kits | Calcium-dependent binding of exosomal membrane externalized PS to Tim4 protein immobilized on magnetic beads/resins; intact exosomes are eluted via calcium chelation with EDTA under mild, non-denaturing conditions | Pan-exosome isolation (no tetraspanin expression bias) with high purity and yield; Mild elution fully preserves exosome structure, surface protein expression, and biological activity; Minimal lipoprotein contamination, superior performance for plasma/serum samples [46]; Magnetic bead format compatible with high-throughput automated processing |
Higher per-sample cost than precipitation or UF isolation methods [46]; Isolation efficiency is highly sensitive to calcium concentration in the sample buffer; Risk of co-isolation of PS-positive apoptotic bodies without proper pre-clearing; Limited commercial options and independent head-to-head validation data in the published literature |
| Multimodal/Combined Isolation Kits | Integrate two complementary isolation principles to balance yield and purity; common combinations include precipitation+SEC, UF+SEC, UF+immunoaffinity, and SEC+immunoaffinity, with a primary concentration step followed by a polishing purification step | Balanced exosome yield and purity, outperforming single-method kits in most matrices; Standardized, integrated workflow eliminates in-house protocol optimization; Optimized for high-contaminant clinical samples (plasma, serum, milk) [48]; Compatible with both high-yield nucleic acid analysis and high-purity proteomics/functional assays |
Longer processing time and more complex workflow than single-method kits; Higher per-sample cost from integrated multi-step consumables; Limited independent validation in published literature for newer products; Optimized performance is often limited to specific validated sample matrices [48] |
Enriching and concentrating on exosomes
In many experiments, an initial isolation step is followed by additional enrichment to reach the purity and concentration needed for specific downstream assays. Combining methods can help balance yield, purity, and practicality.
A common strategy is to use one method for bulk separation and another for polishing. For instance, ultrafiltration can be used to concentrate conditioned medium or plasma before SEC, thereby reducing volume and shortening column runs. Alternatively, some groups pair an initial ultracentrifugation or precipitation step with immunoaffinity capture to enrich for vesicles expressing particular markers while depleting remaining contaminants.
Beyond the choice of separation methods, concentrating EVs to workable volumes is often necessary. Laboratories may use centrifugal filters with defined cut‑offs, vacuum‑based concentrators for relatively clean preparations, or tangential flow filtration for larger volumes that require gentle handling and scalable processing. Each approach requires attention to potential losses, membrane binding, and changes in apparent size distribution, which should be checked by subsequent characterization.
No isolation method is complete without quality control. Characterization confirms that you have enriched for the particle type you expect and helps you understand what else is present.
Nanoparticle tracking analysis (NTA) is now a widely used tool to measure EV size distribution and concentration in solution. NTA tracks the Brownian motion of individual particles in a laser beam to estimate their hydrodynamic diameter and count particles in a defined volume. It can detect size ranges typical of exosomes and microvesicles and, with fluorescent labeling, can focus on vesicles carrying specific markers.
Dynamic light scattering and tunable resistive pulse sensing provide complementary information on particle size and distribution, but they often require cleaner samples and exhibit different sensitivities to polydispersity. In practice, many groups combine particle‑level measurements, such as NTA, with orthogonal techniques to cross‑validate their preparations and to compare isolation conditions.
Protein markers remain central to defining EV preparations. MISEV2018 recommends demonstrating the presence of several positive markers associated with EV membranes and cytosolic components (for example, tetraspanins such as CD9, CD63, and CD81, and cytosolic proteins like TSG101) and the relative absence of markers from non‑vesicular cellular compartments or abundant soluble proteins.
Western blotting or ELISA assays can be used to assess both positive and negative markers, providing a quick readout of enrichment and contamination. Flow cytometry, often using bead‑bound vesicles, extends this approach by enabling multi‑parameter profiling and higher throughput, and immuno‑NTA or related techniques combine marker detection with size and concentration information in a single measurement.
Structural methods add another layer of confirmation. Transmission electron microscopy (TEM) has long been used to visualize EVs, revealing typical cup‑shaped or round vesicle structures and helping to distinguish vesicles from many types of aggregates. Cryo‑electron microscopy can preserve native morphology but is more technically demanding.
Ultimately, many projects also benefit from functional assays that test whether isolated EVs can transfer nucleic acids, proteins, or other signals to recipient cells in biologically relevant ways. While these readouts are context‑dependent, MISEV encourages linking functional data to well‑documented characterization so that observed effects can be attributed to EVs rather than co‑isolated factors.
Even experienced groups encounter issues with exosome experiments. Recognizing common pitfalls early can save time and make your isolation workflows more robust [45].
Low yield is a frequent challenge, especially when working with small sample volumes or low‑secreting cell types. In such cases, increasing starting volume, extending collection time, or choosing methods with higher recovery (even at the cost of purity) may be reasonable if supported by subsequent clean‑up and characterization. Conversely, very high apparent yields with little change in protein content may signal that non‑vesicular material is co‑isolating, especially when using precipitation methods without additional purification.
Contamination with proteins or lipoproteins is another persistent issue, particularly in blood‑derived samples. Including density‑based separation, SEC, or additional washing steps can reduce these contaminants, and checking negative markers as recommended by MISEV2018 helps document success. For all methods, clear reporting of protocols, including rotor types, forces, times, and buffers, improves reproducibility across laboratories.
Standardization efforts such as the MISEV guidelines and related community initiatives provide a framework for documenting and comparing EV studies. Adopting these recommendations—covering sample collection, separation, characterization, functional testing, and reporting—supports more rigorous and comparable work in exosome research. Where possible, using reference materials or participating in inter‑laboratory comparisons can further benchmark performance and highlight areas for protocol improvement.
CUSABIO provides many exosome marker-related proteins and several exosome ELISA kits to help researchers study exosome-associated research.
Table 4. Universal Positive Markers for Exosome Identification
These markers are consistently enriched in exosomes/sEVs across most cell and tissue sources, and are recommended for core exosome characterization by MISEV guidelines.
| Exosome Markers | Core Function in Exosome Biology |
|---|---|
| CD9 | Enriched in exosomal membranes; regulates exosome biogenesis, membrane fusion, and intercellular signal transduction; one of the most widely used pan-exosome markers |
| CD63 | Late endosome/multivesicular body (MVB)-enriched protein; mediates exosomal cargo sorting, exosome biogenesis, and cellular uptake; a canonical marker for endosome-derived exosomes |
| CD81 | Stabilizes exosomal membrane structure via cholesterol binding; modulates exosome secretion, cellular uptake, and immune regulatory functions; a core pan-exosome marker |
| TSG101 | Core regulator of ESCRT-dependent exosome biogenesis; mediates MVB formation and exosomal cargo sorting; a key cytosolic marker for exosome validation |
| PDCD6IP (ALIX) | Modulates both ESCRT-dependent and independent exosome biogenesis via interaction with syntenin-1; regulates MVB budding and exosome release |
| HSPA8 (HSP70) | Abundantly packaged in exosomes; mediates exosomal protein folding, cargo loading, and intercellular stress signal transmission; a validated cytosolic exosome marker |
| HSP90AB1 (HSP90) | Highly enriched in exosomal lumen; regulates exosomal cargo stability and immune cell activation; widely used for exosome characterization |
| FLOT1 (Flotillin-1) | Enriched in exosomal lipid raft microdomains; participates in exosome biogenesis, membrane trafficking, and signal transduction; a conserved exosome membrane marker |
| SDCBP (Syntenin-1) | Key mediator of ALIX-dependent non-ESCRT exosome biogenesis; regulates exosomal cargo sorting and MVB-plasma membrane fusion |
| RAB27A | Master regulator of late-stage exosome trafficking; controls MVB docking to the plasma membrane and calcium-dependent exosome secretion |
Table 5. Cell/Tissue-Specific Exosome Markers
These markers are uniquely enriched in exosomes derived from specific cell types, enabling the identification of exosome tissue origin in biofluids and functional studies.
| Source Cell/Tissue | Exosome Markers | Specificity & Application |
|---|---|---|
| Mesenchymal Stem/Stromal Cells (MSCs) | NT5E (CD73) | Definitive marker for human MSC-derived exosomes; used to validate MSC origin and distinguish from other stromal cell exosomes |
| Mesenchymal Stem/Stromal Cells (MSCs) | THY1 (CD90) | Core MSC surface marker; consistently expressed on MSC-derived exosomes; used for immunocapture of MSC exosomes |
| ENG (CD105) | MSC-specific marker; enriched in MSC exosomes; used for phenotyping and origin validation | |
| Macrophages | CD68 | Pan-macrophage marker; highly enriched in macrophage-derived exosomes; used to trace monocyte/macrophage exosome origin |
| CD14 | Monocyte/macrophage lineage marker; specific for exosomes from classical monocytes and tissue macrophages | |
| T Lymphocytes | CD3D | Pan-T cell marker; exclusively expressed on T cell-derived exosomes; used to identify T cell exosomes in immune studies |
| CD4 | Subset-specific markers for helper T cell (CD4) and cytotoxic T cell (CD8) derived exosomes | |
| B Lymphocytes | CD19 | Pan-B cell marker; enriched in B cell-derived exosomes; used to validate B cell exosome origin |
| HLA-DRA | Highly expressed on B cell exosomes; critical for exosome-mediated antigen presentation | |
| Epithelial Tumor Cells | EPCAM | Tumor exosome marker for epithelial-origin cancers (breast, colorectal, lung); used for liquid biopsy and circulating tumor exosome isolation |
| EGFR | Enriched in exosomes from EGFR-amplified cancers (non-small cell lung cancer, glioblastoma); used for tumor exosome phenotyping | |
| Neurons | L1CAM | Neuron-specific marker; enriched in neuron-derived exosomes (NDEs); the gold standard for NDE isolation from peripheral blood |
| NCAM1 | Pan-neuronal marker; consistently expressed on neuronal exosomes; used for central nervous system exosome origin validation | |
| Vascular Endothelial Cells | PECAM1 (CD31) | Endothelial-specific marker; highly enriched in endothelial cell-derived exosomes; used for vascular exosome studies |
| CDH5 (VE-Cadherin) | Endothelial lineage-specific marker; exclusive to vascular endothelial exosomes; used to assess endothelial exosome cargo | |
| Platelets | ITGA2B | Platelet-specific marker; enriched in platelet-derived exosomes; used to trace platelet exosome origin in thrombosis and inflammation studies |
Getting exosomes is more than just spinning down a sample or adding a kit; it is a series of choices about source, isolation method, enrichment, and quality control that collectively determine what your experiments will see. By aligning those choices with community standards, combining complementary methods when necessary, and building in robust characterization, researchers can obtain small EV preparations that are fit for purpose and reproducible across time and laboratories.
References
[1] G. Raposo, H.W. Nijman, W. Stoorvogel, R. Liejendekker, C.V. Harding, C.J. Melief, H.J. Geuze. B lymphocytes secrete antigen-presenting vesicles [J]. J. Exp. Med. 1996, 183:1161–1172.
[2] H. Valadi, A. Bossios, et al. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells [J]. Nat. Cell Biol. 2007, 9:654–659.
[3] Zong SF, Wang L, et al. Facile detection of tumor-derived exosomes using magnetic nanobeads and SERS nanoprobes [J]. Anal Methods. 2016, 8:5001–08.
[4] Harding CV, Heuser JE, et al. Exosomes: looking back three decades and into the future [J]. J. Cell Biol. 2013, 200:367–371.
[5] B.T. Pan, R.M. Johnstone. Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: selective externalization of the receptor [J]. Cell. 1983, 33:967–978.
[6] Pan BT, Teng K, et al. Electron microscopic evidence for externalization of the transferrin receptor in vesicular form in sheep reticulocytes [J]. J. Cell Biol.1985;101:942–948.
[7] Johnstone RM, Bianchini A, et al. Reticulocyte maturation and exosome release: transferrin receptor containing exosomes shows multiple plasma membrane functions [J]. Blood. 1989, 74:1844–1851.
[8] Li, Q., Ding, Y., et al. (2026). 80 years of extracellular vesicles: From discovery to clinical translation [J]. Extracellular Vesicles and Circulating Nucleic Acids, 7(1), 165.
[9] Omrani, M., Beyrampour-Basmenj, H., et al. (2023). Global trend in exosome isolation and application: An update concept in management of diseases [J]. Molecular and Cellular Biochemistry, 1.
[10] Li, M., Rai, A. J., et al. (2015). An optimized procedure for exosome isolation and analysis using serum samples: Application to cancer biomarker discovery [J]. Methods, 87, 26-30.
[11] Contreras-Naranjo, J. C., Wu, H. J., & Ugaz, V. M. (2017). Microfluidics for exosome isolation and analysis: Enabling liquid biopsy for personalized medicine [J]. Lab on a Chip, 17(21), 3558.
[12] Jiawei, S., Zhi, C., Kewei, T., & Xiaoping, L. (2022). Magnetic bead-based adsorption strategy for exosome isolation [J]. Frontiers in Bioengineering and Biotechnology, 10, 942077.
[13] Xu, K., Jin, Y., Li, Y., Huang, Y., & Zhao, R. (2022). Recent Progress of Exosome Isolation and Peptide Recognition-Guided Strategies for Exosome Research [J]. Frontiers in Chemistry, 10, 844124.
[14] Liu, W. Z., Ma, Z. J., & Kang, X. W. (2022). Current status and outlook of advances in exosome isolation [J]. Analytical and Bioanalytical Chemistry, 414(24), 7123.
[15] Dilsiz, N. (2024). A comprehensive review on recent advances in exosome isolation and characterization: Toward clinical applications [J]. Translational Oncology, 50, 102121.
[16] Wang, J., Ma, P., Kim, D. H., Liu, B. F., & Demirci, U. (2021). Towards Microfluidic-Based Exosome Isolation and Detection for Tumor Therapy [J]. Nano Today, 37, 101066.
[17] Cheng, Y., Qu, X., et al. (2020). Comparison of serum exosome isolation methods on co-precipitated free microRNAs [J]. PeerJ, 8, e9434.
[18] Gao M, Cai J, Zitkovsky HS, Chen B, Guo L. Comparison of Yield, Purity, and Functional Properties of Large-Volume Exosome Isolation Using Ultrafiltration and Polymer-Based Precipitation. Plast Reconstr Surg. 2022 Mar 1;149(3):638-649.
[19] Pölzl L, Nägele F, et al. Exosome Isolation after in vitro Shock Wave Therapy [J]. J Vis Exp. 2020 Sep 10;(163).
[20] Abramowicz, A., Marczak, L., et al. (2018). Harmonization of exosome isolation from culture supernatants for optimized proteomics analysis [J]. PLoS ONE, 13(10), e0205496.
[21] Neerukonda, S. N., Egan, N. A., et al. (2020). A comparison of exosome purification methods using serum of Marek's disease virus (MDV)-vaccinated and -tumor-bearing chickens [J]. Heliyon, 6(12), e05669.
[22] Tangwattanachuleeporn, M., Muanwien, P., Teethaisong, Y., & Somparn, P. (2022). Optimizing Concentration of Polyethelene Glycol for Exosome Isolation from Plasma for Downstream Application [J]. Medicina, 58(11), 1600.
[23] Lobb, R. J., Becker, M., et al. (2015). Optimized exosome isolation protocol for cell culture supernatant and human plasma [J]. Journal of Extracellular Vesicles, 4, 10.3402/jev.v4.27031.
[24] Doyle, L. M., & Wang, M. Z. (2019). Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis [J]. Cells, 8(7), 727.
[25] Zhang, H., & Lyden, D. (2019). Asymmetric flow field-flow fractionation technology for exomere and small extracellular vesicle separation and characterization [J]. Nature Protocols, 14(4), 1027.
[26] Zhang H, Freitas D, Kim HS, et al. Identification of distinct nanoparticles and subsets of extracellular vesicles by asymmetric flow field-flow fractionation [J]. Nat Cell Biol. 2018;20(3):332–343.
[27] Greening DW, Xu R, Ji H, Tauro BJ, Simpson RJ. A protocol for exosome isolation and characterization: evaluation of ultracentrifugation density-gradient separation, and immunoaffinity capture methods [J]. Methods Mol Biol. 2015;1295:179–209.
[28] Iorember FM, Vehaskari VM. Uromodulin: old friend with new roles in health and disease [J]. Pediatr Nephrol. 2014;29(7):1151–1158.
[29] Liu F, Vermesh O, Mani V, Ge TJ, et al. The exosome total isolation chip [J]. ACS Nano. 2017;11(11):10712–10723.
[30] Yang Q, Cheng L, Hu L, et al. An integrative microfluidic device for isolation and ultrasensitive detection of lung cancer-specific exosomes from patient urine [J]. Biosens Bioelectron. 2020;163:112290.
[31] Santana SM, Antonyak MA, Cerione RA, Kirby BJ. Microfluidic isolation of cancer-cell-derived microvesicles from hetergeneous extracellular shed vesicle populations [J]. Biomed Microdevices. 2014;16(6):869–877.
[32] Wunsch BH, Smith JT, et al. Nanoscale lateral displacement arrays for the separation of exosomes and colloids down to 20 nm [J]. Nat Nanotechnol. 2016;11(11):936–940.
[33] Ibsen SD, Wright J, et al. Rapid isolation and detection of exosomes and associated biomarkers from plasma [J]. ACS Nano. 2017;11(7):6641–6651.
[34] Zhao W, Zhang L, et al. Microsphere mediated exosome isolation and ultra-sensitive detection on a dielectrophoresis integrated microfluidic device [J]. Analyst. 2021;146(19):5962–5972.
[35] Wu M, Ouyang Y, Wang Z, et al. Isolation of exosomes from whole blood by integrating acoustics and microfluidics [J]. Proc Natl Acad Sci U S A. 2017;114(40):10584–10589.
[36] Wang Z, Li F, Rufo J, et al. Acoustofluidic salivary exosome isolation: a liquid biopsy compatible approach for human papillomavirus-associated oropharyngeal cancer detection [J]. J Mol Diagn. 2020;22(1):50–59.
[37] Pang B, Zhu Y, Ni J, et al. Extracellular vesicles: the next generation of biomarkers for liquid biopsy-based prostate cancer diagnosis [J]. Theranostics. 2020;10(5):2309–2326.
[38] Wang Z, Wu HJ, et al. Ciliated micropillars for the microfluidic-based isolation of nanoscale lipid vesicles [J]. Lab Chip. 2013;13(15):2879–2882.
[39] Ranjan S, Zeming KK, Jureen R, Fisher D, Zhang Y. DLD pillar shape design for efficient separation of spherical and non-spherical bioparticles [J]. Lab Chip. 2014;14(21):4250–4262.
[40] Zhu H, Lin X, Su Y, Dong H, Wu J. Screen-printed microfluidic dielectrophoresis chip for cell separation [J]. Biosens Bioelectron. 2015;63:371–378.
[41] Aghilinejad A, Aghaamoo M, Chen X, Xu J. Effects of electrothermal vortices on insulator-based dielectrophoresis for circulating tumor cell separation [J]. Electrophoresis. 2018;39(5–6):869–877.
[42] Fraunhofer W, Winter G. The use of asymmetrical flow field-flow fractionation in pharmaceutics and biopharmaceutics [J]. Eur J Pharm Biopharm. 2004;58(2):369–383.
[43] Wu B, Chen X, Wang J, et al. Separation and characterization of extracellular vesicles from human plasma by asymmetrical flow field-flow fractionation [J]. Anal Chim Acta. 2020;1127:234–245.
[44] Kim YB, Yang JS, Lee GB, Moon MH. Evaluation of exosome separation from human serum by frit-inlet asymmetrical flow field-flow fractionation and multiangle light scattering [J]. Anal Chim Acta. 2020;1124:137–145.
[45] Witwer, K. W., Goberdhan, D. C., et al. (2021). Updating MISEV: Evolving the minimal requirements for studies of extracellular vesicles [J]. Journal of Extracellular Vesicles, 10(14), e12182.
[46] Patel GK, Khan MA, et al. Comparative analysis of exosome isolation methods using culture supernatant for optimum yield, purity and downstream applications [J]. Sci Rep. 2019 Mar 29;9(1):5335.
[47] Aliakbari F, Stocek NB, et al. A methodological primer of extracellular vesicles isolation and characterization via different techniques [J]. Biol Methods Protoc. 2024 Feb 13;9(1):bpae009.
[48] Pang B, Zhu Y, et al. Quality Assessment and Comparison of Plasma-Derived Extracellular Vesicles Separated by Three Commercial Kits for Prostate Cancer Diagnosis [J]. Int J Nanomedicine. 2020 Dec 15;15:10241-10256.


Comments
Leave a Comment