Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
| Code | CSB-AP002431HU |
| Abbreviation | Recombinant Human FGF9 protein, partial (Active) |
| MSDS | |
| Size | $142 |
| Order now | |
| Image |
|
| Have Questions? | Leave a Message or Start an on-line Chat |
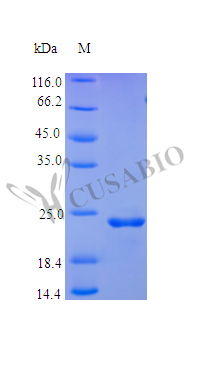
FGF9 plays a critical role in embryonic development, neurogenesis, and tumor progression by signaling through multiple FGF receptors, making precise receptor-binding potency essential for meaningful experimental results. This tag-free recombinant human FGF9 (residues 2–208) demonstrates an ED50 below 0.5 ng/ml in a BaF3 thymidine uptake assay, corresponding to a specific activity exceeding 2.0 × 10⁶ IU/mg — potency that supports use in cell proliferation assays, neuronal and osteogenic differentiation studies, angiogenesis models, and receptor-ligand binding experiments such as SPR or competition ELISA. Produced in *E. coli*, the protein lacks glycosylation, which provides a chemically defined baseline advantageous for quantitative binding kinetics and for use as an ELISA standard or in antibody characterization workflows where post-translational modifications could confound interpretation. Purity exceeding 95% by SDS-PAGE combined with endotoxin levels below 1.0 EU/μg satisfies the criteria typical for both cell-based functional assays and in vivo tumor biology models.
There are currently no reviews for this product.

