Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
Multicellular organisms live in a complex milieu where signaling pathways play a key role in their survival. As important mediators of this signal cascade, tyrosine kinases exert critical effects in various biological processes such as growth, differentiation, metabolism, and apoptosis in response to internal and external stimuli. Among the 90 unique tyrosine kinase genes identified in the human genome, 58 encode receptor tyrosine kinases (RTKs) [1] [2]. Here we provide a brief overview of the RTK superfamily, classification, functions, their regulatory mechanisms in cells, and related diseases.
1. What Are Receptor Tyrosine Kinases?
2. Classification of Receptor Tyrosine Kinases
3. What Do Receptor Tyrosine Kinases Do?
Receptor tyrosine kinases (RTKs) are the largest category of enzyme-linked receptors, which serve as both receptors and enzymes that can bind to ligands and phosphorylate the tyrosine residues of target proteins. RTKs catalyze the transfer of the ATP γ-phosphate to hydroxyl groups of tyrosines on target proteins [3].
RTKs are single-pass membrane proteins composed of three parts: an extracellular domain containing ligand-binding sites, a single α-helix trans-membrane hydrophobic region, and an intracellular region containing a juxtamembrane regulatory portion, a tyrosine kinase catalytic domain (TKD), and a C-terminal tail [4] [5]. The ligand-binding ectodomain differs across RTK subfamilies from Ig-like fold type (VEGFRs), fibronectin type III (FnIII) domains (INSR), and Leucine-rich repeat and cysteine-rich domains (EGFR). The transmembrane domain is an important player in the formation and stabilization of the dimer of the receptor chains. The TKD contains the catalytic sites and is responsible for the catalysis of the ATP-dependent phosphorylation of receptor chains.
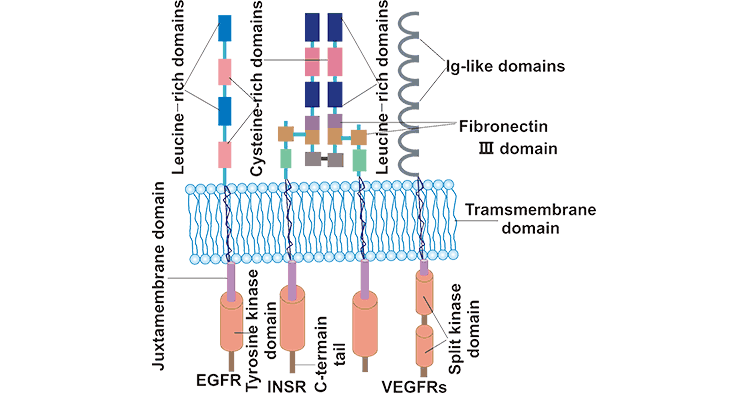
Figure 1: Structure of RTK subfamilies.
*The picture is cites from https://jmhg.springeropen.com/articles/10.1186/s43042-019-0035-0
RTKs are distributed into 19 subfamilies according to their structural and ligand-affinity properties [1] [2] [6]. The LMR1–3 family has recently been revised and removed due to its re-classification as Ser/Thr receptor kinases [32]. The 19 RTK subfamilies include the epidermal growth factor receptors (EGFRs or ErbBs), the fibroblast growth factor receptors (FGFRs), the insulin and the insulin-like growth factor receptors (IR and IGFR), the platelet-derived growth factor receptors (PDGFRs), the vascular endothelial growth factor receptors (VEGFRs), the hepatocyte growth factor receptors (HGFRs), and the nerve growth factor receptors (NGFRs), PTK7/CCK4, Trk, Ror, MuSK, MET, Axl, Tie, EphA/B, Ret, Ryk, DDR, Ros, ALK, and STYK1.
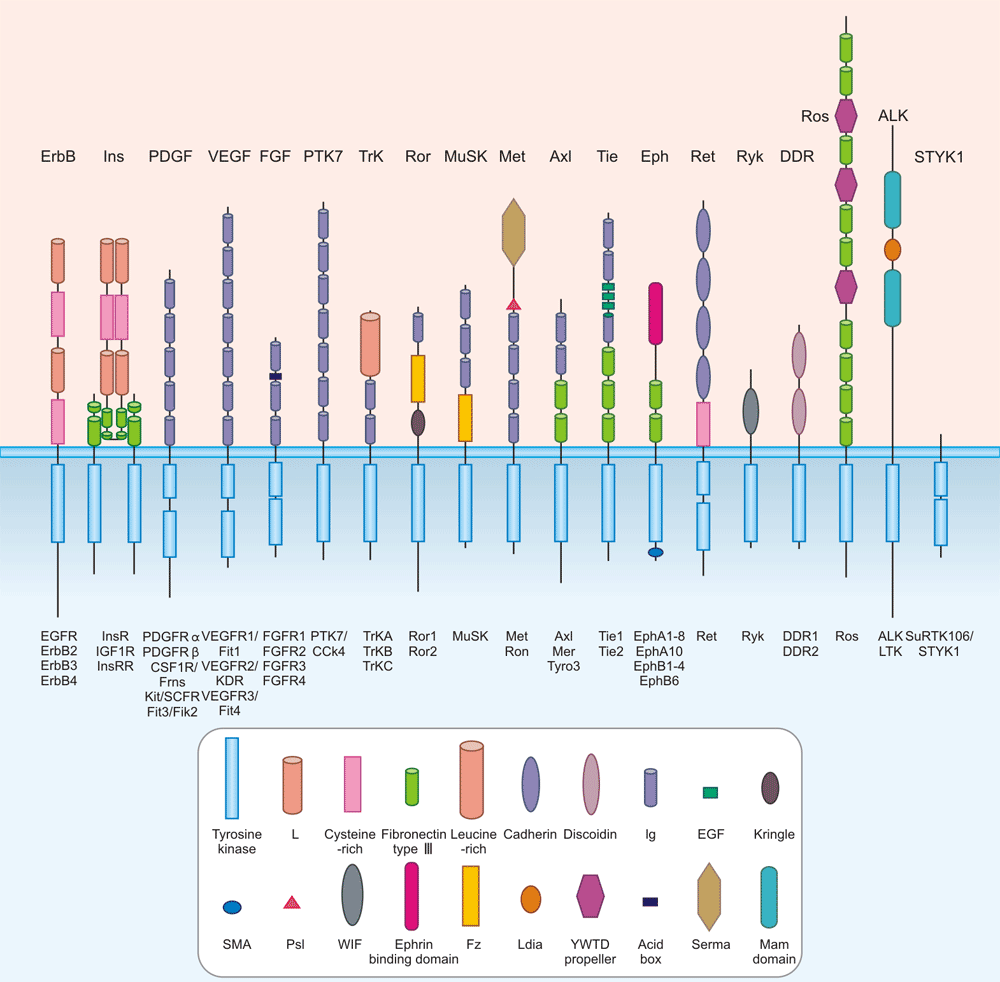
Figure 2: RTK Subfamilies
*The picture is cited from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2914105/
Table: Members of RTK Subfamilies and Their Functions (Cited from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7813211/)
| RTK Subfamilies | Members | Cognate Ligands | Functions |
|---|---|---|---|
| EGFR/ErbB family | EGFR, ErbB2, ErbB3, ErbB4 | EGF, TGFα, EREG, BTC, EPGN, AREG, HB-EGF, NRG1, NRG2, NRG3, NRG4 | Involvement in cell growth and differentiation during embryonic development and in adult life. |
| Insulin receptor family | INSR, IGF1R | Insulin for IR; IGF1 and IGF2 for IGF1R | Promotion of cellular glucose transport, increase of glycogen synthase activity, and regulation of metabolic processes such as protein or lipid synthesis, as well as growth. |
| PDGFR family | PDGFRα, PDGFRβ, CSF1R/Fms, Kit/SCFR, Flt3/Flk2 | PDGF-A, PDGF-B, PDGF-C, PDGF-D, SCF, Flt3L | Modulation of embryonic development, especially the formation of vessels and organs [7]. |
| VEGFR family | VEGFR1, VEGFR2, VEGFR3 | PIGF, VEGFB, VEGFC, VEGFD | Regulation of the angiogenesis development and the vascular system. |
| FGFR family | FGFR1, FGFR2, FGFR3, FGFR4 | FGF1, FGF2, FGF3, FGF4, FGF5, FGF6, FGF7, FGF8, FGF9, FGF10, FGF11, FGF12, FGF13, FGF14, FGF16, FGF17, FGF18, FGF20, FGF22 | Induction of cell growth, differentiation, migration and chemotaxis, angiogenesis, and cell survival. |
| PTK7 family | PTK7/CCK4 | CCK and gastrin | Involvement in the gastrointestinal ( gastric acid secretion, pancreatic enzyme release, gallbladder emptying, gut motility, and energy homeostasis) and neural (regulates appetite, pain, anxiety, and wakefulness) physiology. |
| NGFR/Trk family | TRKA, TRKB, TRKC | NGF, BDNF, NT-3, and NT-4 | Involvement in neuronal survival and differentiation in the developing and mature nervous system [8]. |
| HGFR/Met family | Met, RON | HGF/SF, MSP | Mediation of embryogenesis, tissue regeneration, wound healing, as well as the formation of nerve and muscle in normal cells [9]. |
| EPHR family | EPHA1, EPHA2, EPHA3, EPHA4, EPHA5, EPHA6, EPHA7, EPHA8, EPHA10, EPHB1, EPHB2, EPHB3, EPHB4, EPHB6 | Ephrin A1, Ephrin A2, Ephrin A3, Ephrin A4, Ephrin A5, Ephrin B1, Ephrin B2, Ephrin B3, Ephrin B4, Ephrin B5 | Ephrin-EPHR signaling is implicated in the activation, migration, adhesion, and proliferation of immune cells [27]. |
| AXL/TAM family | AXL, MER, TYRO3 | GAS6 appears to bind all three TAMs, with the highest affinity for AXL, PROS1 predominantly binds TYRO3 and MER [25]. | TAM receptors play no essential role in embryonic development but are involved in phagocytic clearance of apoptotic cells in adult organisms [26]. |
| TIE receptor family | Tie1, Tie2 | Ang1, Ang2, Ang3, Ang4 | Tie1 is required for normal embryonic vascular development. When Tie2 binds with its ligand, it can promote the cleavage of vascular basement membrane, the activation, migration and proliferation of vascular endothelial cells, and the formation of new capillaries. |
| RYK family | RYK | Wnt1, Wnt3a | Control of aspects of neuronal differentiation in the developing brain [10]. |
| DDR family | DDR1, DDR2 | Collagen type I, Collagen type II, Collagen type III, Collagen type IV, Collagen type X | DDR1 is essential for mammary gland development [28] and DDR2 is important for the growth of long bones [29]. |
| RET receptor family | RET (RET9, RET43, and RET51) | GFLs : GDNF, neurturin (NRTN), artemin (ARTN), and persephin (PSPN) | Involvement in cell proliferation, differentiation, and survival. |
| ROS receptor family | ROS | Not known ligands | The ectopic expression and the production of variable mutant forms of ROS kinase has been reported in a number of cancers, such as glioblastoma multiforme, and non-small cell lung cancer. |
| LTK receptor family | LTK, ALK | ALKAL1, ALKAL2 | Protein fusion, over-expression, and activating mutations of ALK is implicated in various types of cancer. |
| ROR family | ROR1, ROR2 | Wnt5A, Wnt5B, and Wnt16 [30] | Regulation of development of the axial and paraxial mesoderm, the nervous system and the neural crest, the axial and appendicular skeleton, and the kidney, and other embryonic developmental processes. |
| MuSK receptor family | MUSK | LRP4 | Involvement in the formation and stabilization of neuromuscular junctions (NMJs) [31]. |
| STYK1 family | RTK106/STYK1 | Not known ligands | Involvement in tumor progression and migration. |
RTKs serve as regulators involved in many important cellular processes, including cell-to-cell communication, cell proliferation, differentiation, survival, motility, morphogenesis, metabolism, and cell cycle control [11] [12]. Several studies have shown that some RTKs are detected in the nucleus and that these nuclear-localized RTKs participate in transcriptional regulation, DNA replication, DNA damage response, and drug resistance [13].
RTKs exist as monomers (except for IR and IGFR) before binding to signal molecules and keep inactive. When a ligand binds to the extracellular domain of the RTK, two monomer RTK molecules dimerize on the membrane and the tails of the intracellular domains of the two receptors contact each other [11]. The trans-autophosphorylation between the dimer pair destabilizes cis-autoinhibition, activating RTK catalytic activity [14] [15]. Autophosphorylation of the TKD produces phosphorylated tyrosine site chains, which function as binding sites for intracellular signaling proteins containing phosphotyrosine-recognition domains, such as the SH2 domain and PTB domain [16] [17]. Docking of protein substrates evokes a series of biochemical reactions in cells through several different signal transduction pathways, including RAS/MAPK, PI-3 K/AKT, and JAK2/STAT signaling pathways.
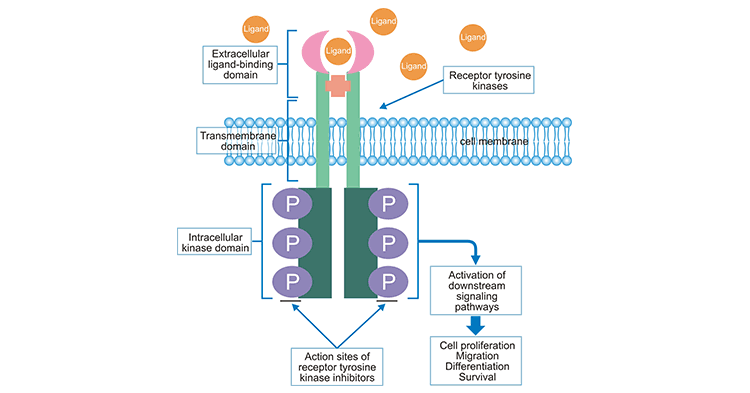
Figure 3: RTK Signaling Pathway
*The picture is cited form https://www.frontiersin.org/articles/10.3389/fonc.2020.01642/full
RTKs play an important role during development and throughout life as described above. It is not surprising that they are implicated in various developmental diseases and malignancies. Genetic mutations, overexpression, or dysregulation of RTKs have been associated with a wide range of human diseases, including cancer, diabetes, inflammation, severe bone disorders, and arteriosclerosis. Anomalous activation of RTKs has been linked to the initiation, progression, and prognosis of several human malignancies. Studies have shown that about 30% of RTKs are mutated or overexpressed in various human cancers [18]. Overexpression of ErbB-1 and ErbB-2 has been detected in breast [19] [20], lung [21], and prostate [22] [23] cancer and is related to significant aggressiveness and poor prognosis.
Abnormalities in RTKs' activation or signaling lead to many pathologies, especially cancers, pushing the development of various drugs that block RTK signaling. Some drugs have been successfully applied for the treatment of many types of cancer [24]. EGFR signal transduction pathway plays an important role in the cell proliferation, injury repair, invasion, and neovascularization of tumor cells. EGFR targeting drugs have become a new hotspot in tumor therapy in recent years. These drugs are mainly divided into two categories: small-molecule tyrosine kinase inhibitors (TKI) that act on the intracellular region of receptors such as gefitinib and erlotinib, and monoclonal antibodies that function on the extracellular region of receptors, including cetuximab, ABX-EGF and EMD 72000.
References
[1] D R Robinson, Y M Wu, et al. The protein tyrosine kinase family of the human genome [J]. Oncogene. 2000 Nov 20; 19(49):5548-57.
[2] Manning G, Plowman GD, et al. Evolution of protein kinase signaling from yeast to man [J]. Trends Biochem Sci 2002, 27:514–520.
[3] Hunter T. The Croonian lecture, 1997. The phosphorylation of proteins on tyrosine its role in cell growth and disease [J]. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 1998; 353: 583-605.
[4] Hubbard SR. Structural analysis of receptor tyrosine kinases [J]. Prog Biophys Mol Biol. 1999;71:343–58.
[5] Li E, Hristova K. Receptor tyrosine kinase transmembrane domains: function, dimer structure and dimerization energetics [J]. Cell Adhes Migr 2010, 4(2):249–254.
[6] Manning G, Whyte DB, et al. The protein kinase complement of the human genome [J]. Science. 2002;298:1912–34.
[7] Andrae J, Gallini R, Betsholtz C. Role of platelet-derived growth factors in physiology and medicine [J]. Genes Dev. 2008;22:1276–1312.
[8] Reichardt LF. Neurotrophin-regulated signalling pathways [J]. Philos Trans R Soc Lond B Biol Sci 2006, 361: 1545–1564.
[9] Zhang, Y., Xia, M., Jin, K. et al. Function of the c-Met receptor tyrosine kinase in carcinogenesis and associated therapeutic opportunities [J]. Mol Cancer 17, 45 (2018).
[10] Lyu J, Yamamoto V, Lu W. Cleavage of the Wnt receptor Ryk regulates neuronal differentiation during cortical neurogenesis [J]. Dev Cell 2008, 15: 773–780.
[11] Zhenfang Du & Christine M. Lovly. Mechanisms of receptor tyrosine kinase activation in cancer [J]. Molecular Cancer volume 17, Article number: 58 (2018).
[12] Christine Chiasson-MacKenzie and Andrea I McClatchey. Cell-Cell Contact and Receptor Tyrosine Kinase Signaling [J]. Cold Spring Harb Perspect Biol. 2018 Jun 1;10(6):a029215.
[13] Du Y., Hsu J.L., et al. Nuclear Functions of Receptor Tyrosine Kinases. In: Wheeler D., Yarden Y. (eds) Receptor Tyrosine Kinases: Structure, Functions and Role in Human Disease [J]. Springer, New York, NY. 2015.
[14] Fantl WJ, Johnson DE, Williams LT. Signaling by Receptor Tyrosine Kinases [J]. Annu Rev Biochem. 1993;62:453–481.
[15] Lemmon MA, Schlessinger J. Cell signaling by receptor tyrosine kinases [J]. Cell. 2010;141:1117–34.
[16] Huse M and Kuriyan J. The conformational plasticity of protein kinases [J]. Cell, 2002, 109:275–282.
[17] Schlessinger J. Cell signaling by receptor tyrosine kinase [J]s. Cell. 2000;103:211–25.
[18] Lu Z., Jiang G., Jensen P., Hunter T. Epidermal growth factor-induced tumor cell invasion and metastasis initiated by dephosphorylation and downregulation of focal adhesion kinase [J]. Mol Cell Biol. 2001;21:4016–4031.
[19] Bose R., Kavuri S.M., et al. Activating HER2 mutations in HER2 gene amplification negative breast cancer [J]. Cancer Discov. 2013;3:224–237.
[20] Nakajima H., Ishikawa Y., et al. Protein expression, gene amplification, and mutational analysis of EGFR in triple-negative breast cancer [J]. Breast Cancer. 2014;21:66–74.
[21] Lynch T.J., Bell D.W., et al. Activating mutations in the epidermal growth factor receptor underlying responsiveness of non-small-cell lung cancer to gefitinib [J]. N Engl J Med. 2004;350:2129–2139.
[22] Peraldo-Neia C., Migliardi G., et al. Epidermal growth factor receptor (EGFR) mutation analysis, gene expression profiling and EGFR protein expression in primary prostate cancer [J]. BMC Cancer. 2011;11:31.
[23] Fu M., Zhang W., et al. Mutation status of somatic EGFR and KRAS genes in Chinese patients with prostate cancer (PCa) Virchows Arch [J]. 2014;464:575–581.
[24] Joseph Schlessinger. Receptor Tyrosine Kinases: Legacy of the First Two Decades [J]. Cold Spring Harb Perspect Biol. 2014 Mar; 6(3): a008912.
[25] Tsou WI, Nguyen KQ, et al. Receptor tyrosine kinases, TYRO3, AXL, and MER, demonstrate distinct patterns and complex regulation of ligand-induced activation [J]. J Biol Chem 2014, 289: 25750–25763.
[26] Lemke G. Biology of the TAM receptors [J]. Cold Spring Harb Perspect Biol2013, 5: a009076.
[27] Thayer K. Darling and Tracey J. Lamb. Emerging Roles for Eph Receptors and Ephrin Ligands in Immunity [J]. Front. Immunol., 04 July 2019.
[28] Vogel, W. F., Aszódi, A., et al. Discoidin domain receptor 1 tyrosine kinase has an essential role in mammary gland development [J]. Mol. Cell. Biol. 2001, 21, 2906–2917.
[29]Labrador, J. P., Azcoitia, V., Tuckermann, et al. The collagen receptor DDR2 regulates proliferation and its elimination leads to dwarfism [J]. EMBO Rep.2001, 2, 446–452.
[30] Kamizaki K, Endo M, et al. Role of Noncanonical Wnt Ligands and Ror-family Receptor Tyrosine Kinases in the Development, Regeneration, and Diseases of the Musculoskeletal System [J]. Dev Dyn (2021) 250(1):27–38.
[31] Stevan R. Hubbard and Kavitha Gnanasambandan. Structure and Activation of MuSK, a Receptor Tyrosine Kinase Central to Neuromuscular Junction Formation [J]. Biochim Biophys Acta. 2013 Oct; 1834(10): 2166–2169.
[32] Wendler F. The LMTK-family of kinases: Emerging important players in cell physiology and disease pathogenesis [J]. Biochimica et Biophysica Acta (BBA) - Molecular Basis of Disease 2018.


Comments
Leave a Comment