
Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
| Code | CSB-RA826096A0HU |
| Size | US$210 |
| Order now | |
| Image |
|
| Have Questions? | Leave a Message or Start an on-line Chat |
| Application | Recommended Dilution |
|---|---|
| WB | 1:500-1:5000 |
| IHC | 1:50-1:200 |
| IF | 1:20-1:200 |
| FC | 1:20-1:200 |
E2F1 is a pivotal transcription factor that orchestrates cell cycle progression by regulating genes essential for DNA replication and S-phase entry. Its interaction with the retinoblastoma protein places it at a critical node in tumor suppressor pathways, making E2F1 dysregulation a hallmark of numerous malignancies. For researchers investigating cell proliferation, apoptosis, or oncogenic transformation, reliable detection of this transcription factor is fundamental to understanding disease mechanisms.
This recombinant monoclonal antibody, generated against a synthetic peptide derived from human E2F1, offers the consistency and reproducibility that demanding experimental workflows require. As a sequence-defined recombinant clone, it eliminates the lot-to-lot variability inherent in traditional hybridoma-derived antibodies, ensuring that your results remain comparable across extended studies and collaborative projects.
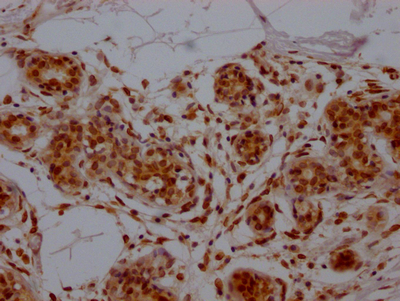
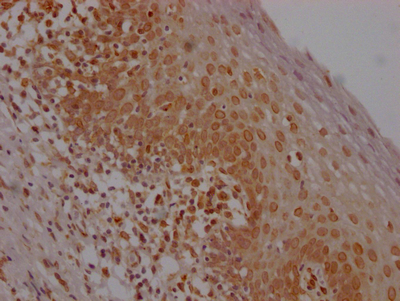
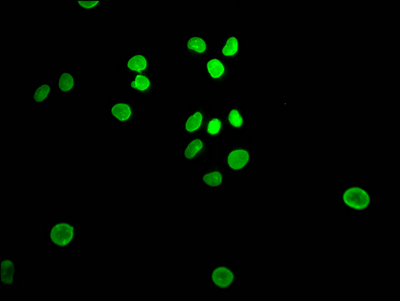
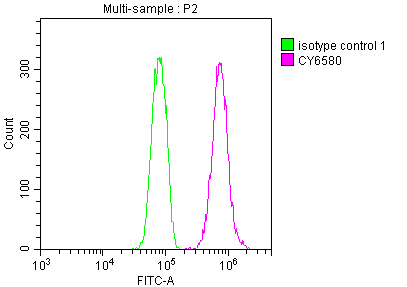
Validation data demonstrates robust performance across multiple platforms. Western blot analysis detects E2F1 in Jurkat and PC-3 whole cell lysates as well as rat heart tissue, with an observed band at approximately 60 kDa rather than the predicted 47 kDa—a shift commonly attributed to post-translational modifications such as phosphorylation or glycosylation that regulate E2F1 activity. Immunohistochemistry staining has been validated in paraffin-embedded human breast cancer and tonsil tissues, while immunofluorescence studies in HeLa cells confirm clear nuclear localization consistent with E2F1's role as a transcription factor. Flow cytometry applications have also been validated using HeLa cells, providing flexibility for quantitative single-cell analysis.
With confirmed cross-species reactivity in human and rat samples, this antibody serves researchers exploring epigenetic regulation, nuclear signaling cascades, and cancer biology across diverse experimental models and methodological approaches.
There are currently no reviews for this product.

