Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
For a researcher or clinician facing a tumor sample, a critical question looms: how fast is this cancer actually growing? Accurately measuring cancer growth rate is more than an academic exercise; it shapes diagnosis, prognosis, and therapeutic strategy. The key to unlocking this answer lies in understanding the cell cycle, and specifically, the proteins that serve as its phase-specific indicators: cell cycle markers. These molecular beacons, from the well-known Ki-67 to the phase-specific cyclins, have evolved from fundamental discoveries in cell biology to non-negotiable tools in modern oncology.
This article will guide you through their core biology, the new technologies used to detect them, and their pivotal role in diagnosing, stratifying, and treating cancer today.
Table of Contents
2. What Are Cell Cycle Markers?
3. Cell Cycle Markers in Oncology
4. Emerging Tools and Technologies Used to Detect Cell Cycle Markers
5. Future Directions and Research Frontiers of Cell Cycle Markers
The cell cycle is a meticulously ordered series of events leading to cell growth, DNA replication, and division into two daughter cells.
The eukaryotic cell cycle is usually divided into four primary stages:
A fifth state, G0, represents a quiescent or differentiated state outside the active cycle.
Progression through these phases is controlled by cyclin‑dependent kinases (CDKs), which are activated by cyclins and inhibited by CDK inhibitors (CKIs) such as p21 and p27. When this control system is intact, cells divide only when they should; when it is broken, uncontrolled proliferation and cancer can follow [1].
Cell cycle markers are proteins whose expression levels or activity fluctuate in a periodic manner throughout the cell cycle. They serve as essential tools for identifying the current phase of a cell (G1, S, G2, or M), monitoring cell cycle progression, and understanding dysregulation of proliferation in diseases such as cancer [2]. These markers are crucial for both basic biological research and clinical applications, including cancer diagnosis, prognosis, and the development of targeted therapies [3].
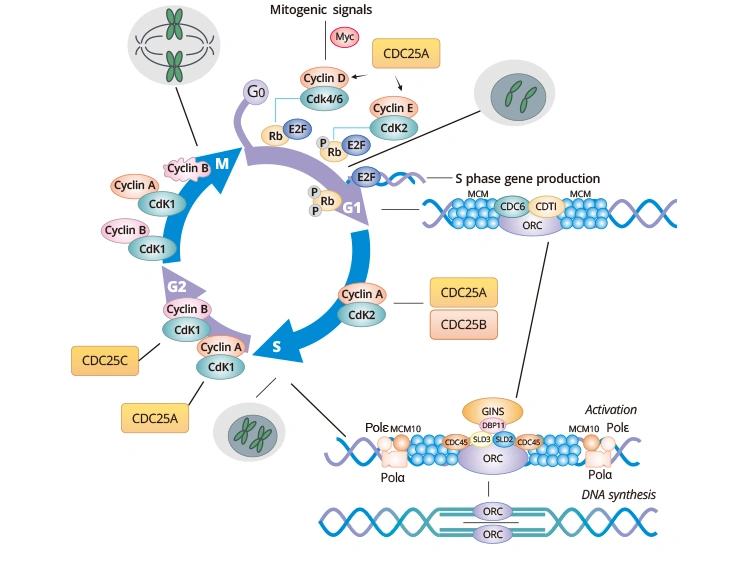
Figure. Cell cycle markers at each phase of the cell cycle
Cell cycle markers can be categorized based on their functional roles and the specific phase of the cell cycle they indicate.
The G1 phase is a period of growth and preparation for DNA synthesis. Key regulators here determine whether a cell commits to division or exits to G0.
This checkpoint represents the irreversible commitment to DNA replication.
During G2, the cell prepares for mitosis, ensuring DNA replication is complete and undamaged.
Mitosis involves chromosome segregation and cell division.
These markers are expressed in all active phases of the cycle (G1, S, G2, M) but not in G0, serving as general indicators of proliferative activity.
In oncology, cell cycle markers also influence diagnosis, prognosis, and treatment choices.
Cancer often involves sustained proliferative signaling and disrupted cell‑cycle checkpoints. Amplification or overexpression of cyclins, constitutive CDK activity, and loss of CKIs or p53‑pathway components are common events in many tumor types. These alterations not only drive growth but also change the landscape of cell‑cycle markers, typically increasing proliferation markers and shifting phase distributions [26].
Understanding these patterns can highlight vulnerabilities. Tumors with strong cyclin D–CDK4/6 dependency, for example, may respond to CDK4/6 inhibitors, and changes in proliferation markers during treatment can act as pharmacodynamic readouts [26].
Ki‑67 has become a workhorse proliferation biomarker in cancer. In invasive breast cancer, the Ki‑67 labeling index is widely studied as a prognostic factor and is used in some guidelines to help stratify risk and support treatment decisions. High Ki‑67 often correlates with higher grade, increased metastatic potential, and poorer outcomes, although cutoffs and scoring methods vary between studies and centers [27-29].
Beyond breast cancer, Ki‑67 is used in grading neuroendocrine tumors and explored in many other malignancies. Reviews emphasize its clinical utility but also highlight pitfalls such as variability in antibodies, staining protocols, scoring, and intratumor heterogeneity, all of which need standardization for reliable use [27-29].
Cyclin D1 overexpression, often due to gene amplification, is a classic example of a cell‑cycle regulator with biomarker potential, especially in certain lymphomas and breast cancers. CDKs themselves, particularly CDK4/6, are now therapeutic targets, and ongoing studies examine whether expression of these kinases or their downstream effects can predict response or resistance to inhibitors [26].
CKIs such as p16 are used diagnostically in some settings. For example, p16 immunostaining is a surrogate marker for oncogenic HPV in cervical and oropharyngeal lesions. Patterns of p21 and p27 expression have also been linked to prognosis in various cancers, reflecting their role as brakes on CDK activity [26,30].
As research methods evolve, the study of cell cycle markers has expanded far beyond traditional microscopy. The new frontier focuses on real-time, high-dimensional insight into how cells behave and diversify.
Recent technologies are enabling the dynamic tracking to cell cycle markers within living systems.
These methods are transforming the cell cycle from a static concept to a dynamic system observed in real time.
Integrated "omics" approaches—combining proteomics, transcriptomics, and metabolomics—have deepened our understanding of cell cycle networks. Computational models now predict how signaling perturbations alter proliferation patterns.
Such strategies help researchers identify novel, context-specific markers, especially in cancer subtypes where regulatory circuits are rewired.
Looking ahead, cell cycle research is converging with computational power and personalized medicine, promising even deeper insights into proliferation and control.
Artificial intelligence (AI) now plays a growing role in pattern recognition across imaging, genomic, and proteomic datasets. Algorithms can detect subtle variations in marker expression invisible to traditional analysis, accelerating biomarker validation pipelines [32].
In precision oncology, the goal is to tailor therapies to each tumor’s proliferation signature. Cell cycle markers inform predictive models of drug response—helping clinicians distinguish patients who may benefit most from CDK inhibitors or DNA repair-targeting drugs.
This integration of molecular insight with clinical need marks the next evolution of translational cell biology.
Mounting studies have focused on cell cycle markers due to their important roles in assessing tumor progression and prognosis. Demand for products related to cell cycle markers has also increased. CUSABIO has been committed to providing high-quality scientific research products for the majority of scholars and scientific institutions. Here is a list of cell cycle marker-associated products.
| Cell Cycle Markers | Cell Cycle Markers | Cell Cycle Markers | Cell Cycle Markers |
|---|---|---|---|
| E2F transcription factors | E2F1 | p16(INK4a) | p16(INK4a) |
| E2F2 | p15(INK4b) | p15(INK4b) | |
| E2F3 | p21 | p21 | |
| E2F4 | p27 | p27 | |
| E2F5 | ATM | ATM | |
| E2F6 | Chk2 | Chk2 | |
| E2F7 | Ki-67 | Ki-67 | |
| E2F8 | MCM | MCM2 | |
| Cyclins | cyclin A1 | MCM3 | |
| cyclin A2 | MCM4 | ||
| cyclin B1 | MCM5 | ||
| cyclin B2 | MCM6 | ||
| cyclin B3 | MCM7 | ||
| cyclin D1 | MCM10 | ||
| cyclin D2 | Cdc25 | Cdc25A | |
| cyclin D3 | Cdc25B | ||
| cyclin E1 | Cdc25C | ||
| cyclin E2 | ORC | ORC1 | |
| cyclin E | ORC2 | ||
| Cdks | Cdk1 | ORC3 | |
| Cdk2 | ORC4 | ||
| Cdk4 | ORC5 | ||
| Cdk6 | ORC6 | ||
| PLK1 | PLK1 | Cdc6 | Cdc6 |
| CDCA | CDCA1/NUF1 | Cdc45 | Cdc45 |
| CDCA2 | Cdt1 | Cdt1 | |
| CDCA3 | Geminin | Geminin | |
| CDCA4 | PCNA | PCNA | |
| CDCA5 | Aurora A | Aurora A | |
| CDCA6 | Aurora B | Aurora B | |
| CDCA7 | H3S10PH | H3S10PH | |
| CDCA8 | PLK1 | PLK1 |
From foundational regulators like cyclins to clinical indicators such as Ki-67, cell cycle markers continue to bridge basic biology and medicine. They allow researchers to visualize growth, understand pathology, and forecast therapeutic outcomes.
As technology advances and interdisciplinary approaches expand, the study of cell cycle markers is poised to yield not only deeper biological understanding but also tangible clinical advances. The cell cycle, once viewed as a series of static steps, is now appreciated as a dynamic landscape that continues to generate critical insights into cancer and beyond.
References
[1] Pellarin, I., Favero, A., et al. (2025). Cyclin-dependent protein kinases and cell cycle regulation in biology and disease [J]. Signal Transduction and Targeted Therapy, 10, 11.
[2] ZHONG, J., LIU, J., et al. (2025). Cell cycle proteins: Linking the cell cycle to tumors [J]. Oncology Research, 33(6), 1335.
[3] Otto, T., & Sicinski, P. (2017). Cell cycle proteins as promising targets in cancer therapy [J]. Nature Reviews. Cancer, 17(2), 93.
[4] Michalides R. Prognosis for G1 cell-cycle regulators: useful for predicting course of disease and for assessment of therapy in cancer [J]. J Pathol. 1999 Aug;188(4):341-3.
[5] Swellam, M., El-Arab, L. R. E., & Adly, A. (2008). Prognostic value of cell-cycle regulators and cellular biomarkers in laryngeal squamous cell carcinoma [J]. Clinical Biochemistry, 41(13), 1059-1066.
[6] Mineo, T. C., Ambrogi, V., Mineo, D., & Baldi, A. (2005). Long-term disease-free survival of patients with radically resected thymomas [J]. Cancer, 104(10), 2063-2071.
[7] Bogdanovic Lj, Radojevic-Skodric S, et al. Immunohistochemical detection of cyclin E in transitional cell carcinoma [J]. J BUON. 2011 Oct-Dec;16(4):727-32.
[8] Michalides, R., Balkenende, A., et al. (2002). Cyclin A is a prognostic indicator in early stage breast cancer with and without tamoxifen treatment [J]. British Journal of Cancer, 86(3), 402.
[9] Hashiguchi Y, Tsuda H, et al. Relationship between HPV typing and the status of G2 cell cycle regulators in cervical neoplasia [J]. Oncol Rep. 2004 Sep;12(3):587-91.
[10] Huang, J. X., Yan, W., Song, et al. (2005). Relationship between proliferative activity of cancer cells and clinicopathological factors in patients with esophageal squamous cell carcinoma [J]. World Journal of Gastroenterology: WJG, 11(19), 2956.
[11] Wang, J., Liu, X., Chu, H., & Chen, J. (2022). Cell division cycle associated 2 (CDCA2) upregulation promotes the progression of hepatocellular carcinoma in a p53-dependant manner [J]. PeerJ, 10, e13535.
[12] Cui, H., Peng, J., Li, Z., Lyu, J., Zhu, F., & Qin, H. (2021). Cell division cycle associated 8: A novel diagnostic and prognostic biomarker for hepatocellular carcinoma [J]. Journal of Cellular and Molecular Medicine, 25(24), 11097.
[13] DeLuca JG, Howell BJ, et al. Nuf2 and Hec1 Are Required for Retention of the Checkpoint Proteins Mad1 and Mad2 to Kinetochores [J]. Curr Biol (2003) 13(23):2103–9.
[14] Thadani R, Uhlmann F, Heeger S. Condensin, Chromatin Crossbarring and Chromosome Condensation [J]. Curr Biol (2012) 22(23): R1012–21.
[15] Peng A, Lewellyn AL, Schiemann WP, Maller JL. Repo-Man Controls a Protein Phosphatase 1-Dependent Threshold for DNA Damage Checkpoint Activation [J]. Curr Biol (2010) 20(5):387–96.
[16] Vagnarelli P. Repo-Man at the Intersection of Chromatin Remodelling, DNA Repair, Nuclear Envelope Organization, and Cancer Progression [J]. Adv Exp Med Biol (2014) 773:401–14.
[17] Ayad NG, Rankin S, et al. Tome-1, a Trigger of Mitotic Entry, Is Degraded During G1 via the APC [J]. Cell (2003) 113(1):101–13.
[18] Yoshida K. Cell-Cycle-Dependent Regulation of the Human and Mouse Tome-1 Promoters [J]. FEBS Lett (2005) 579(6):1488–92.
[19] Wang L, Zhu G, Yang D, Li Q, Li Y, Xu X, et al. The Spindle Function of CDCA4 [J]. Cell Motil Cytoskeleton (2008) 65(7):581–93.
[20] Ladurner R, Kreidl E, et al. Sororin Actively Maintains Sister Chromatid Cohesion [J]. EMBO J (2016) 35(6):635–53.
[21] Watrin E, Demidova M, Watrin T, Hu Z, Prigent C. Sororin Pre-mRNA Splicing Is Required for Proper Sister Chromatid Cohesion in Human Cells [J]. EMBO Rep (2014) 15(9):948–55.
[22] Vincenz C, Kerppola TK. Different Polycomb Group CBX Family Proteins Associate With Distinct Regions of Chromatin Using Nonhomologous Protein Sequences [J]. Proc Natl Acad Sci USA (2008) 105(43):16572–7.
[23] Prescott JE, Osthus RC, et al. A Novel C-Myc-Responsive Gene, JPO1, Participates in Neoplastic Transformation [J]. J Biol Chem (2001) 276(51):48276–84.
[24] Thijssen PE, Ito Y, Grillo G, et al. Mutations in CDCA7 and HELLS Cause Immunodeficiency-Centromeric Instability-Facial Anomalies Syndrome [J]. Nat Commun (2015) 6:7870.
[25] Jenness C, Giunta S, et al. HELLS and CDCA7 Comprise a Bipartite Nucleosome Remodeling Complex Defective in ICF Syndrome [J]. Proc Natl Acad Sci USA (2018) 115(5): E876–85.
[26] Pellarin, I., Favero, A., et al. (2025). Cyclin-dependent protein kinases and cell cycle regulation in biology and disease [J]. Signal Transduction and Targeted Therapy, 10, 11.
[27] Menon SS, Guruvayoorappan C, Sakthivel KM, Rasmi RR. Ki-67 protein as a tumour proliferation marker [J]. Clin Chim Acta. 2019 Apr;491:39-45.
[28] Davey MG, Hynes SO, Kerin MJ, Miller N, Lowery AJ. Ki-67 as a Prognostic Biomarker in Invasive Breast Cancer [J]. Cancers (Basel). 2021 Sep 3;13(17):4455.
[29] Li LT, Jiang G, Chen Q, Zheng JN. Ki67 is a promising molecular target in the diagnosis of cancer (review) [J]. Mol Med Rep. 2015 Mar;11(3):1566-72.
[30] Sun, X., Bizhanova, A., et al. (2017). Ki-67 Contributes to Normal Cell Cycle Progression and Inactive X Heterochromatin in p21 Checkpoint-Proficient Human Cells [J]. Molecular and Cellular Biology, 37(17), e00569-16.
[31] Buettner F, Natarajan KN, et al. Computational analysis of cell-to-cell heterogeneity in single-cell RNA-sequencing data reveals hidden subpopulations of cells [J]. Nat Biotechnol. 2015 Feb;33(2):155-60.
[32] Esteva, A., Robicquet, A., Ramsundar, B. et al. A guide to deep learning in healthcare [J]. Nat Med 25, 24–29 (2019).
[33] Cosetta Bertoli, Jan M. Skotheim, et al. Control of cell cycle transcription during G1 and S phases [J]. Nat Rev Mol Cell Biol. 2013 Aug; 14(8): 518–528.
[34] Malysa, A., Zhang, X. M., & Bepler, G. (2024). Minichromosome Maintenance Proteins: From DNA Replication to the DNA Damage Response. Cells, 14(1), 12.
[35] Kulkarni AA, Loddo M, et al. DNA replication licensing factors and aurora kinases are linked to aneuploidy and clinical outcome in epithelial ovarian carcinoma [J]. Clin Cancer Res 2007, 13: 6153–6161.


Comments
Leave a Comment