Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
CADM1 is a cell adhesion molecule that belongs to the immunoglobulin superfamily, and it has a ubiquitous presence in the human body. Apart from its abundance in multiple normal tissues where it serves various functions, CADM1 is also detected on numerous tumor cells. Notably, CADM1 acts as a Tumor Suppressor Gene (TSG) in the growth, infiltration, and metastasis of several types of cancer. Several studies demonstrate that in many malignant tumors, methylation of the CADM1 promoter reduces or deletes its expression, leading to widespread cancer cell infiltration and metastasis.
Furthermore, the ability of CADM1 to suppress cell metastasis may be linked to T-cell-mediated cellular immunity, specifically through CD8+ T-cells. These discoveries offer novel insights for cancer treatment. Being a part of the immunoglobulin superfamily, CADM1 presents a promising target for tumor diagnosis and the investigation of new tumor immunotherapies.
The Immunoglobulin superfamily (Ig SF) is renowned for being the most diverse group of Cell Adhesion Molecules (CAMs). Its members include major histocompatibility complex class I and II molecules, T-cell receptor complex proteins, viral receptors, and cell surface glycoproteins [1-2]. These adhesion molecules contain an immunoglobulin (Ig)-like structural domain in their structure and do not depend on Ca2+ [3-4].
Immunoglobulin superfamily adhesion molecules are widely expressed in endothelial cells, epithelial cells, leukocytes, and the nervous system. Several Ig SF members have been identified as biomarkers of cancer progression. For instance, MCAM, L1CAM (CD171), NCAM (CD56), PECAM-1 (CD31), ALCAM (CD166), and ICAM-1 (CD54) are linked with metastatic progression in different types of cancers such as melanoma, glioma, breast, ovarian, endometrial, prostate and colon cancers [5-6].
Cell adhesion molecule 1 (CADM1), also known as Necl2, TSLC1, IGSF4, RA175, and Syn CAM, belongs to the immunoglobulin superfamily of cell adhesion molecules. The protein encoded by CADM1 has three distinct regions: extracellular, transmembrane, and cytoplasmic. The extracellular region comprises 422 amino acids and forms three immunoglobulin C2-type structural domains via disulfide bonds. The transmembrane region is an α-helical hydrophobic structure, while the intracellular region has 46 amino acid residues.
The cytoplasmic region is similar in structure to blood type glycoprotein c and features a FERM protein binding motif adjacent to the transmembrane region. Additionally, there is a PDZ binding motif at the pegylated terminus that allows it to bind to the corresponding protein and perform its specific functions (Figure 1) [8-9].
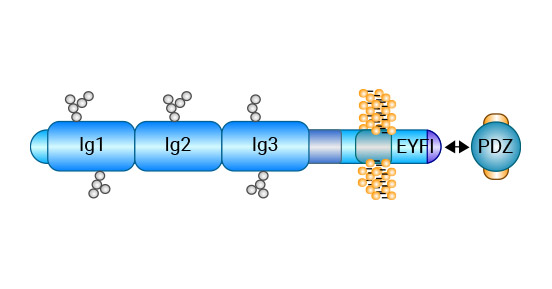
Figure 1. CADM1 Structure [9]
CADM1 is expressed in various tissues including the brain, testis, lung, and epithelial tissues. This protein plays a crucial role in mediating cell adhesion between adjacent cells by facilitating homogeneous transfer interactions. Additionally, CADM1 interacts with other proteins such as actin-binding protein 4.1B/DAL-1 and different scaffolding proteins like membrane-associated guanylate kinases (i.e., MPP1, MPP2, MPP3, CASK, and Pals2), to participate in various biological processes. Recent studies have identified CADM1 as a tumor suppressor gene with significant potential in controlling excessive cell proliferation and suppressing tumor invasion and metastasis. Thus, targeting CADM1 may prove beneficial in developing new treatments for cancer [10-11].
CADM1 is a recently discovered TSG that plays critical roles in various cellular processes, including intercellular adhesion, cell motility, signal transduction, and immune regulation. Despite considerable research efforts aimed at investigating the tumor suppression functions of this gene, its specific in vivo function and molecular mechanism of tumor suppression remain unclear. Further studies are needed to elucidate these aspects of CADM1 biology.
Methylation-induced inactivation of tumor suppressor genes is a key early mechanism implicated in tumorigenesis. CADM1 expression deletion, caused by promoter methylation, has been closely linked to the development of various cancers including liver, lung, intestinal, gastric, prostate, ovarian, cervical, breast, pancreatic, nasopharyngeal, glioma, and acute leukemia.
In hepatocellular carcinoma, several key proteins involved in cell cycle regulation and growth (P15, P16, P21, P27, Cyclin D1, Cyclin D3, CDK4, CDK6, phopho-Rb, Rb, and E2F1) were studied. Western Blot experiments demonstrated that CADM1 overexpression upregulated the expression of the critical cell cycle protein Rb. Rb is a crucial tumor suppressor as it inhibits the activity of the transcription factor E2F, thereby preventing cells from entering S-phase and keeping them dormant during G0/G1 phase, leading to stunted cell growth. Consequently, CADM1 may hinder tumor cell growth via the Rb-E2F pathway [12-13].
In ovarian cancer, the upregulation of CADM1 leads to a concomitant increase in IFI44L and C4BPA expression. Additionally, CADM1 overexpression inhibits the activation of PI3K/Akt/mTOR pathway by activating LXR/RXR signaling while suppressing GF-1 and FXYD2 expression. The inhibition of PI3K/Akt/mTOR pathway upstream activation ultimately leads to the downregulation of APP, EDN1, and Rapla, as well as the upregulation of TGFBI. Moreover, the activation of ROS/JNK pathway induces the upregulation of apoptosis-related proteins caspase-3 and caspase-8, resulting in apoptosis in ovarian cancer cells. Furthermore, TGFBI and ROS/JNK pathways inhibit proliferation and metastasis of ovarian cancer cells (Figure 2) [14].
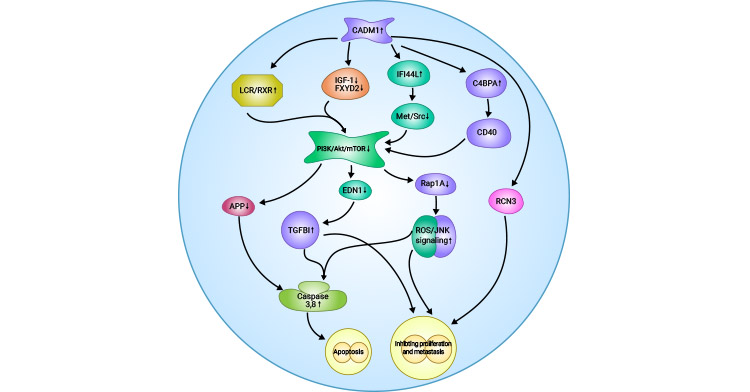
Figure 2. Possible mechanism of CADM1 inhibition in ovarian cancer [14]
A different study found that the tumor suppression provided by CADM1 is reliant on immune surveillance mediated by T cells. Specifically, activated natural killer (NK) cells and CD8+ T cells were observed to impede tumor growth through CRTAM cellular expression. Furthermore, the interaction between CRTAM and CADM1 led to cytotoxic effects and increased γ-interferon secretion by CD8+ T cells. In vivo experiments demonstrated that the CRTAM-CADM1 interaction promotes the rejection of CADM1-expressing tumor cells by NK cells, resulting in their elimination (Figure 3) [15].
Injection of tumor cells expressing CADM1 into the peritoneal cavity of nude mice at an early stage results in efficient killing of the tumor cells by NK cells [15]. These observations support the hypothesis that CADM1 functions as a tumor antigen molecule and its down-regulation may enable tumor cells to evade immune surveillance conducted by the host.
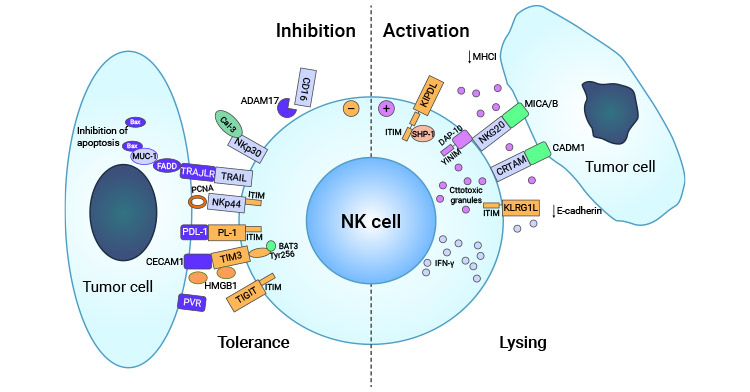
Figure 3. CRTAM-CADM1 is involved in tumor immune escape mechanism [15]
The CADM1 gene was originally identified as a tumor suppressor in non-small cell lung cancer (NSCLC) and its oncogenic potential was later confirmed in nude mice [16]. Furthermore, CADM1 was found to interact with HER2 and integrin α6β4, forming a complex that inhibits the proliferation and invasion of lung squamous cell carcinoma by disrupting the STAT3 activity (Figure 4) [17].
In the case of lung adenocarcinoma, CADM1 methylation was associated with smoking-related factors, leading to the hypothesis that smoking may contribute to gene methylation, indirectly implicating smoking in the development of lung cancer. Additional investigations demonstrated that CADM1 suppresses the proliferation of lung adenocarcinoma cells via interactions with cytoplasmic band 4.1 and MAGu K protein [18].
Furthermore, a recombinant virus known as Ad sp-E1A(Δ24)-TSLC1 (CADM1) was generated through the integration of Ad sp-E1A(Δ24) into a dual-regulatory Ad sp-E1A(Δ24) vector. The efficacy of ad sp-E1A(Δ24)-TSLC1 was observed to be dose-dependent against lung cancer cells. Overexpression of CADM1 in these cells resulted in substantial apoptosis and increased activation of caspase-8, caspase-3, and PARP. Remarkably, intratumoral injection of Ad-sp-E1A(Δ24)-TSLC 1 (CADM1) in a nude mouse transplantation tumor model led to a significant suppression in tumor volume and improved survival rates [19].
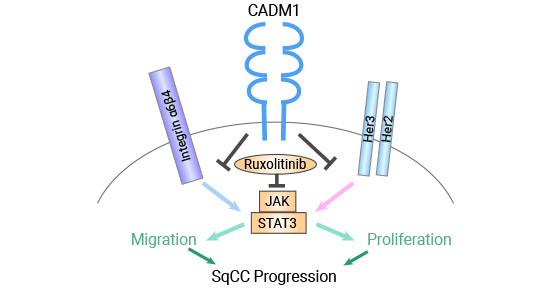
Figure 4. CADM1 inhibits squamous cell carcinoma by decreasing STAT3 activity [17]
Research has indicated that CADM1 can form a direct interaction with DAL-1/4.1B proteins through the FERM structural domain, working synergistically to suppress tumor invasion and metastasis in colorectal cancer. Notably, investigations using DAL-1/4.1B knockout mice revealed that CADM1 distribution remained unaffected by the absence of DAL-1/4.1B, whereas deletion of the FERM binding domain within CADM1 led to alterations in DAL-1/4.1B expression on the cell membrane [20-22].
CADM1 and DAL-1/4.1B play crucial roles in the cytoskeletal cascade by forming stable adhesions between adjacent cells, with actin being a key component of these tissues. Inactivation of CADM1 leads to a decrease in the expression of DAL-1/4.1B and MPP3, which adversely impacts the stability of cell membranes and skeletal adhesion and junctions that are implicated in the development, progression, and metastasis of colorectal cancer. Additionally, in colon adenocarcinoma, CADM1 enhances integrin α6β4 interactions, thereby promoting cellular stability and reducing tumor cell motility [21-22].
Studies have shown that in cervical cancer cell lines, the CADM1 promoter is hypermethylated in 9 out of 11 (82%). Notably, CADM1 gene silencing via promoter hypermethylation is a frequent occurrence that contributes to the progression from high-risk HPV and high-risk cervical intraepithelial neoplasia (CIN) lesions to invasive cervical cancer [23-24].
Within hepatocellular carcinoma, patients with CADM1 promoter region methylation exhibited markedly higher postoperative tumor recurrence rates in comparison to individuals lacking promoter region methylation [25]. Meanwhile, ovarian cancer research has indicated that CADM1 inhibits proliferation and metastasis of ovarian cancer cells while promoting apoptosis via inhibition of PI3K/Akt/mTOR activation [26]. Additionally, CADM1 has been demonstrated to interact with Tiam1 in T-cell leukemia, leading to the human T-cell leukemia virus type I transformation and invasion of surrounding tissues by adult T-cell leukemia cells (Figure 5) [27].
Melanoma studies have indicated that CADM1 overexpression suppresses MMP2 and MMP9 expression, contributing to its inhibitory effects [28]. In immortalized kidney cells, CADM1 binds to HER3, leading to reduced cell proliferation [29-30]. Furthermore, CADM1 has been identified as a candidate for regulating the progression of squamous epithelial carcinoma, with patients displaying tumors lacking CADM1 expression exhibiting poorer prognoses than those with positive expression [31].
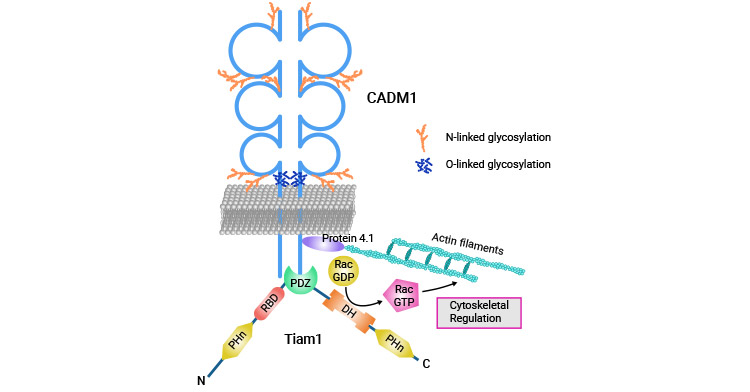
Figure 5. CADM1 interacts with Tiam1 to promote human T-cell leukemia [27]
CADM1 is a recently discovered anti-oncogene whose deletion is closely linked to promoter methylation. Detection of such methylation serves as an effective biomarker for tumor diagnosis and prognosis monitoring. Additionally, CADM1 plays a pivotal role in tumor immunosurveillance, and its low expression or deletion is strongly correlated with tumor immune evasion. Ongoing development and testing of drugs targeting CADM1, including recombinant viral Ad sp-E1A(Δ24)-TSLC1 (CADM1), which promotes apoptosis of tumor cells and viral particle propagation in tumor tissues, is already underway. With the potential to enhance CADM1 function, this gene shows great promise for clinical drug applications in oncology as a promising tumor suppressor gene.
To fully support researchers and pharmaceutical companies in their research on CADM1 in cancers, CUSABIO presents CADM1 active protein (CSB-MP004425MO) to support your research on the mechanism of CADM1 or its potential clinical value (click for the full list of CADM1 products: CADM1 Proteins; CADM1 antibodies).
CADM1 protein:
Recombinant Mouse Cell adhesion molecule 1(Cadm1),partial (Active)
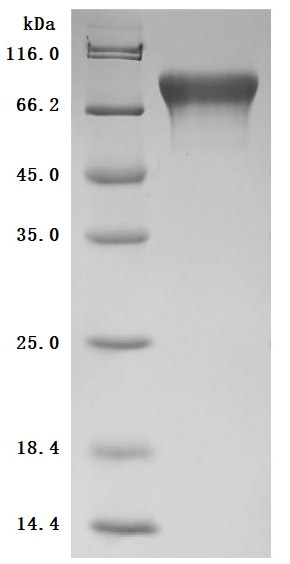
The purity was greater than 98.6% as determined by SDS-PAGE.(Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
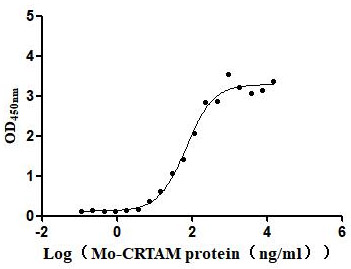
Immobilized Mouse CADM1 at 2μg/mL can bind Mouse CRTAM (CSB-MP614820MO), the EC50 is 52.61-92.92 ng/mL.
References
[1] Verschueren, Erik, et al. "The immunoglobulin superfamily receptome defines cancer-relevant networks associated with clinical outcome." Cell 182.2 (2020): 329-344.
[2] Huene, Aidan L., et al. "A family of unusual immunoglobulin superfamily genes in an invertebrate histocompatibility complex." Proceedings of the National Academy of Sciences 119.40 (2022): e2207374119.
[3] Soltantoyeh, Tahereh, et al. "Chimeric antigen receptor (CAR) T cell therapy for metastatic melanoma: challenges and road ahead." Cells 10.6 (2021). 1450.
[4] von Lersner, Ariana, Lenny Droesen, and Andries Zijlstra. "Modulation of cell adhesion and migration through regulation of the immunoglobulin superfamily member ALCAM/CD166." clinical & experimental metastasis 36.2 (2019): 87-95.
[5] Velychko, L. M., et al. "Expression of lymphocyte activation markers CD 54 (ICAM-1), CD 5, CD 95 (FAS) and neutrophil activation marker CD15 in the peripheral blood of patients with intermediate uveitis and healthy individuals." (2021).
[6] Wai Wong, Chee, Danielle E. Dye, and Deirdre R. Coombe. "The role of immunoglobulin superfamily cell adhesion molecules in cancer metastasis." International journal of cell biology 2012 (2012).
[7] Kasai, Yutaka, et al. "Trans-homophilic interaction of CADM1 promotes organ infiltration of T-cell lymphoma by adhesion to vascular endothelium." Cancer science 113.5 (2022): 1669-1678.
[8] Funaki, Toko, et al. "CADM1 promotes malignant features of small-cell lung cancer by recruiting 4.1 R to the plasma membrane." Biochemical and Biophysical Research Communications 534 (2021): 172-178.
[9] Salie, Muneeb. Investigating candidate genes identified by genome-wide studies of granulomatous diseases in susceptibility to tuberculosis: ANXA11 and the CADM family. Diss. Stellenbosch: University of Stellenbosch, 2010.
[10] Wikman, Harriet, et al. "Loss of CADM1 expression is associated with poor prognosis and brain metastasis in breast cancer patients." oncotarget 5.10 ( 2014): 3076.
[11] Sakurai-Yageta, Mika, et al. "Tumor suppressor CADM1 is involved in epithelial cell structure." Biochemical and biophysical research communications 390.3 (2009): 977-982.
[12] Watanabe, Toshiki. "Adult T-cell leukemia: molecular basis for clonal expansion and transformation of HTLV-1-infected T cells. "Blood. The Journal of the American Society of Hematology 129.9 (2017): 1071-1081.
[13] Zhang, Wu, et al. "CADM1 regelates the G1/S transition and represses tumorigenicity through the Rb-E2F pathway in hepatocellular carcinoma." Hepatobiliary & Pancreatic Diseases International 15.3 (2016): 289-296.
[14] Si, Xiaoqiang, et al. "CADM1 inhibits ovarian cancer cell proliferation and migration by potentially regulating the PI3K/Akt/mTOR pathway." Biomedicine & Pharmacotherapy 123 (2020): 109717.
[15] Vuletić, Ana, et al. "Cross-talk between tumor cells undergoing epithelial to mesenchymal transition and natural killer cells in tumor microenvironment in colorectal cancer." Frontiers in Cell and Developmental Biology (2021): 3107.
[16] Kuramochi, Masami, et al. "TSLC1 is a tumor-suppressor gene in human non-small-cell lung cancer." Nature genetics 27.4 (2001): 427-430.
[17] Vallath, Sabari, et al. "CADM1 inhibits squamous cell carcinoma progression by reducing STAT3 activity." Scientific Reports 6.1 (2016): 1-12.
[18] Murakami, Shigefumi, et al. "Trans-homophilic interaction of CADM1 activates PI3K by forming a complex with MAGuK-family proteins MPP3 and Dlg." PLoS One 9.2 (2014): e82894.
[19] Lei, Wen, et al. "Tumor suppressor in lung cancer-1 (TSLC1) mediated by dual-regulated oncolytic adenovirus exerts specific antitumor actions in a mouse model." Acta Pharmacologica Sinica 34.4 (2013): 531-540.
[20] Ru Zhang, Zhenglei Xu, and Qinghong Tan. "Expression of CADM1 and DAL-1/4.1 B in Colorectal Cancer. "Chinese Journal of Gerontology 14 (2014): 3845-3847 .
[21] Beaulieu, Jean-François. "Integrin α6β4 in colorectal cancer: expression, regulation, functional alterations and use as a biomarker. "Cancers 12.1 ( 2019): 41.
[22] Cai, Qian, Anding Zhu, and Li Gong. "Exosomes of glioma cells deliver miR-148a to promote proliferation and metastasis of glioblastoma via targeting CADM1." Bulletin du cancer 105.7-8 (2018): 643-651.
[23] Hamashima, Chisato. "Emerging technologies for cervical cancer screening." Japanese Journal of Clinical Oncology 51.9 (2021): 1462-1470.
[24] van Zummeren, Marjolein, et al. "HPV E4 expression and DNA hypermethylation of CADM1, MAL, and miR124-2 genes in cervical cancer and precursor lesions." Modern Pathology 31.12 (2018): 1842-1850.
[25] Wu, Dong-Mei, et al. "Down-regulated lncRNA DLX6-AS1 inhibits tumorigenesis through STAT3 signaling pathway by suppressing CADM1 promoter methylation in liver cancer stem cells." Journal of Experimental & Clinical Cancer Research 38 (2019): 1-17.
[26] Si, Xiaoqiang, et al. "CADM1 inhibits ovarian cancer cell proliferation and migration by potentially regulating the PI3K/Akt/mTOR pathway." Biomedicine & Pharmacotherapy 123 (2020): 109717.
[27] Masuda, Mari, et al. "CADM1 interacts with Tiam1 and promotes invasive phenotype of human T-cell leukemia virus type I-transformed cells and adult T- cell leukemia cells." Journal of Biological Chemistry 285.20 (2010): 15511-15522.
[28] You, Yan, et al. "CADM1/TSLC1 inhibits melanoma cell line A375 invasion through the suppression of matrix metalloproteinases." Molecular Medicine Reports 10.5 (2014): 2621-2626.
[29] Sarkar, Bidhan, et al. "Degradation of p47 by autophagy contributes to CADM1 overexpression in ATLL cells through the activation of NF-κB." Scientific Reports 9.1 (2019): 1-14.
[30] Böhm, Allan, et al. "Molecular mechanisms, diagnostic aspects and therapeutic opportunities of micro ribonucleic acids in atrial fibrillation." International Journal of Molecular Sciences 21.8 (2020): 2742.
[31] Saito-Sasaki, Natsuko, et al. "Cell Adhesion Molecule 1 (CADM1) Is an Independent Prognostic Factor in Patients with Cutaneous Squamous Cell Carcinoma ." Diagnostics 11.5 (2021): 830.


Comments
Leave a Comment