Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
| Code | CSB-E04609m |
| Size | 96T,5×96T,10×96T |
| Price | Request a Quote |
| Trial Size |
24T ELISA Kit Trial Size (Only USD$150/ kit) * Sample kit cost can be deducted as a $30 credit for each 96-assay kit of the same analyte and brand you subsequently purchase within six months until depleted. More details >> Interested in a trial size? Please leave a message below.
|
| Have Questions? | Leave a Message or Start an on-line Chat |
| Intra-assay Precision (Precision within an assay): CV%<8% | ||||||
| Three samples of known concentration were tested twenty times on one plate to assess. | ||||||
| Inter-assay Precision (Precision between assays): CV%<10% | ||||||
| Three samples of known concentration were tested in twenty assays to assess. | ||||||
| To assess the linearity of the assay, samples were spiked with high concentrations of mouse IL-18 in various matrices and diluted with the Sample Diluent to produce samples with values within the dynamic range of the assay. | ||||||
| Sample | Serum(n=4) | |||||
| 1:1 | Average % | 88 | ||||
| Range % | 82-94 | |||||
| 1:2 | Average % | 97 | ||||
| Range % | 91-103 | |||||
| 1:4 | Average % | 93 | ||||
| Range % | 88-99 | |||||
| 1:8 | Average % | 94 | ||||
| Range % | 90-100 | |||||
| The recovery of mouse IL-18 spiked to levels throughout the range of the assay in various matrices was evaluated. Samples were diluted prior to assay as directed in the Sample Preparation section. | ||||||
| Sample Type | Average % Recovery | Range | ||||
| Serum (n=5) | 95 | 90-100 | ||||
| EDTA plasma (n=4) | 99 | 94-107 | ||||
| These standard curves are provided for demonstration only. A standard curve should be generated for each set of samples assayed. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
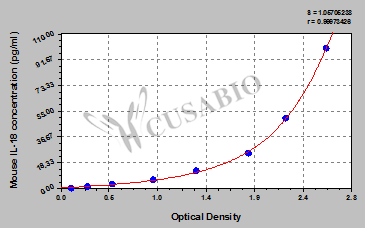
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
This mouse IL-18 ELISA kit uses the quantitative sandwich enzyme immunoassay technique to measure the levels of mouse IL-18 in the samples, including serum, plasma, cell culture supernates, and tissue homogenates. Antibody specific for IL-18 has been pre-coated onto the microplate. Standards and samples are pipetted into the wells and any IL-18 present is bound by the immobilized antibody. After removing any unbound substances, a biotin-conjugated IL-18 antibody is added to the wells. After washing, avidin conjugated Horseradish Peroxidase (HRP) is added to the wells, forming an antibody-antigen-enzyme-labeled antibody complex. Following a wash to remove any unbound HRP-avidin, the TMB substrate solution is added to the wells and the color develops into blue. The color changes from blue to yellow after adding the stop solution to the wells. The color intensity is proportional to the amount of IL-18 bound in the initial step.
IL-18 is a powerful pro-inflammatory cytokine involved in the regulation of both innate and acquired immune responses. It plays a key role in autoimmune, inflammatory, and infectious diseases. IL‐18 binds to IL‐18Rα and IL‐18Rβ, activating NF‐κB and inducing inflammatory mediators such as adhesion molecules, chemokines, and cellular adhesion molecules. In certain cell types, IL18 also induces MAPK and PI3K/ATK signaling pathways resulting in the generation and release of pro-inflammatory cytokines. IL-18 is associated with obesity, insulin resistance, hypertension, and dyslipidemia. High expression of IL-18 has been found in many cancer types, including colorectal cancer, multiple myeloma, and pancreatic cancer, and has been associated with cell migration, invasion, proliferation, and metastasis of tumor cells.
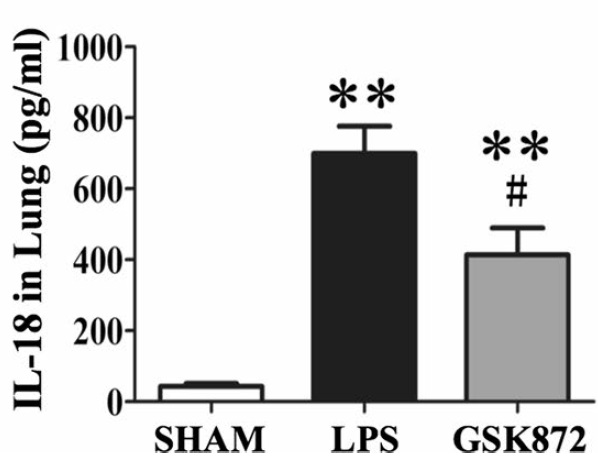
Sample type: Tissue homogenate(Lung tissues)
Sample species: Mouse
Review: IL-1β and IL-18 level in lung tissues measured by ELISA. Each bar represents mean ± SEM (n = 6). *p < 0.05, **p < 0.01 versus sham mice; #p < 0.05, ##p < 0.01 versus LPS-induced lung injury mice.
By Anonymous
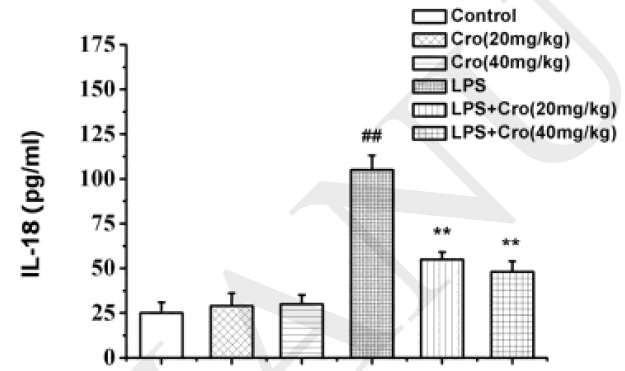
Sample type: Cell culture supernatant
Sample species: Mouse
Review: ELISA was used to determine (A) the levels of IL-1β, (B) IL-18, and (C) TNFα in the hippocampus. ELISA showed that the level of IL-1β, IL-18, and TNFα in the hippocampus on 24h after LPS injection was significantly increased in the LPS group, but reversed in the LPS mice pretreated with Crocin, Data are presented as mean ± SEM, n = 10 mice/group, ##p < 0.01 versus the control group, *P<0.05 and **P<0.01 versus the LPS group.
By Anonymous
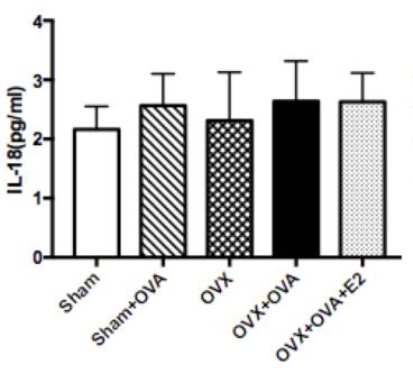
Sample type: Biological fluids(Bronchoalveolar lavage fluid)
Sample species: Mouse
Review: Productions of IL-6, IL-1 β, IL-18, and TNF- α in BALF. Data were presented as means ± SEM (n = 6 in each group). *P < 0.05 **P < 0.001. ***P < 0.001.
By Anonymous
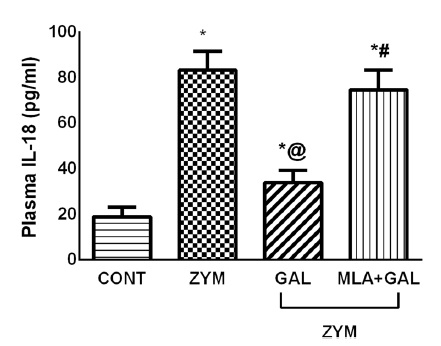
Sample type: Plasma (anticoagulant)
Sample species: Mouse
Review: Effect of GAL (4 mg/kg; i.p) and/or MLA (5.6 mg/kg; i.p) on plasma levels of [a] Cr, [b] Cys-C and [c] IL-18, as well as renal [d] NGAL content in ZYM-induced AKI. Data are expressed as the mean of 6 mice ± SD. Statistical analysis was performed using a one-way analysis of variance (ANOVA) followed by Tukey’s Multiple Comparison test; as compared to CONT(*), ZYM (@), and ZYM + GAL (#) groups, P < 0.05. GAL was injected 1 and 6 h post-ZYM challenge, while MLA was administered 15 min before GAL. AKI: acute kidney injury; CONT: control; Cr: creatinine; Cys-C: cystatin C; GAL: galantamine; MLA: methyllycaconitine, IL-18: interleukin-18; NGAL: neutrophil gelatinase-associated lipocalin; ZYM: zymosan.
By Anonymous
Sample type: Serum
Sample species: Mouse
Sample dilution: No dilution
Review: I have detected influenza virus infection in the peripheral blood supernatant of mice using CSB-E04609m. The operation is simple, convenient, and easy to get started, with very high sensitivity and very accurate detection results.
By Anonymous

Sample type: Plasma (anticoagulant)
Sample species: Mouse
Sample dilution: No dilution
Review: The CSB-E04609m IL18ELISA was used to detect irradiated mouse plasma and the OD value was 0.129.
By Anonymous

