Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
Transferrin receptor TFR1 is one of the key members mediating the entry of iron ions into cellular channels and plays a critical role in regulating cellular iron metabolism and maintaining iron homeostasis. Cancer cells require large amounts of iron for rapid proliferation, leading to a significant upregulation of cell surface transferrin receptor 1 (TfR1), which mediates iron uptake by binding to the iron-carrying protein transferrin. Capitalizing on this phenomenon and the rapid endocytosis rate of TfR1, Xin Zhou's group at the Dana-Farber Cancer Institute in the United States developed the transferrin receptor-targeted chimera (TransTAC), a heterodimeric specific antibody model for membrane protein degradation [1]. TransTAC is designed to drive coendocytosis of target proteins of interest with TfR1 from the cell surface, and to drive the target protein into the lysosomal degradation pathway. This study was published in Nature on September 25, 2024, and TransTAC represents a promising new class of bifunctional antibody family that can be used to precisely modulate membrane proteins and target cancer therapies.
Existing studies have confirmed that TFR1 is highly expressed in many tumor cells and is a potential tumor marker, and that treatment targeting TFR1 can effectively inhibit tumor growth and metastasis. In addition, TFR1 is also associated with other diseases such as anemia and iron metabolism disorders. Therefore, therapeutic strategies targeting TFR1 to regulate intracellular iron levels could play an important role in the clinical application of related diseases.
2. How's the Mechanism of Action of TFR1?
Transferrin receptor protein 1 (TFR1) is a crucial type II transmembrane protein that regulates intracellular iron transport across the membrane. TFR1 comprises of two homodimeric subunits, which are linked by disulfide bonds. Each monomer consists of a short N-terminal region, a single transmembrane region, and a large extracellular C-terminal region. This C-terminal region, serving as an additional functional region, contains the same protein present in transferrin (Tf) (Figure 1) [2-4]. At present, two similar transferrin receptors, TFR1 and TFR2, have been identified. Under normal physiological conditions, TFR1 interacts with Tf to facilitate iron uptake. This bound form is the main way of iron presence in the blood [5-6].
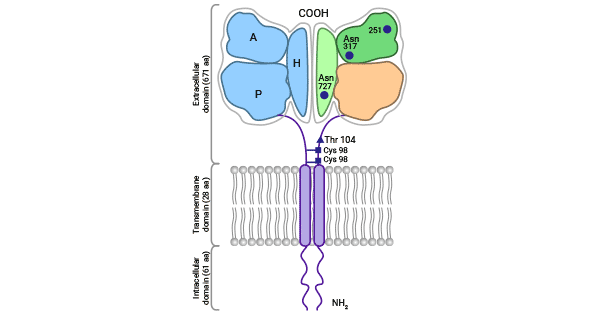
Figure 1. TFR1 Structure [2]
TFR1, a membrane protein expressed across various cell and tissue types in the human body, plays a crucial role in facilitating iron transport and metabolism. Its extensive presence includes immune system, hematopoietic system (e.g., bone marrow stem cells, red blood cells, and leukocytes), nervous system (e.g., neurons and glial cells), reproductive system, heart, liver, and kidney. TFR1 expression levels are susceptible to various factors such as intracellular iron concentration, cellular differentiation status, hormonal regulation, and inflammatory status [6-8].
TFR1's crucial physiological function is to bind with transferrin (Tf), thus facilitating the cellular uptake of iron by endocytosis. This Tf-TFR1 system is a vital pathway for the body to acquire iron ions. The process involves the binding of Tf and iron (Fe3+ or Fe2+) and a subsequent structural change that results in encapsulation of iron ions within the protein, forming Tf-Fe2+. At physiological pH, TFR1 binds to Tf-Fe2+, and the Tf-TFR1 complex undergoes clathrin-mediated endocytosis (Figure 2) [10-12].
The intracellular Tf-TFR1 complex then proceeds to the endosome for acidification, where amino acid residues interact, causing conformational changes that lead to the release of iron ions. TFR1 is recycled to the cell surface through the Golgi complex, completing the transport of iron ions. In summary, TFR1 plays an integral role in maintaining cellular and tissue functions. Regulating TFR1 to balance intracellular iron concentration and ensure iron homeostasis is essential for optimal physiological functioning [10-12].
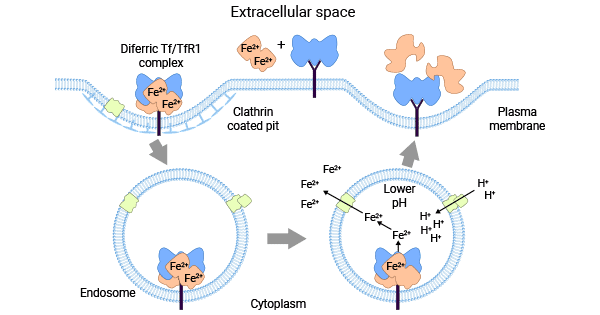
Figure 2. Tf-TFR1 system balances intracellular iron concentration [2]
Abnormal or reduced TFR1 expression can lead to cellular iron deficiency, whereas excess iron may catalyze the production of reactive oxygen species (ROSs) and cause biomolecule damage. Cells have developed various mechanisms to regulate TFR1 expression to ensure adequate iron while avoiding its toxicity. However, the exact mechanisms and roles of aberrant TFR1 expression in disease are incompletely understood.
Different factors may affect the expression and function of TFR1 in cells. At the transcriptional level, hypoxia-inducible factor (HIF), c-Myc, GATA1, Ets-1, erythropoietin Stat5, and other transcription factors can regulate TFR1 expression. At the post-transcriptional level, IRP1 and IRP2 binding to IRE in TFR1 mRNA are key determinants [13-15].
Meanwhile, CD133 (PROM1) acts as a negative regulator. EGF receptor, c-Abl, and MARCH8 also play a role at the post-translational level. A study revealed that FLCN can control TFR1 expression, a protein involved in iron metabolism. This control occurs during or after protein synthesis by helping the Tf-TFR1 complex interact with Rab11-bound endosomes, which circulate within the cell and return to the cell membrane (Figure 3) [13-17].
TFR1 plays a role in regulating various diseases. In glioma, TFR1 affects multiple mechanisms related to inflammation, DNA, and cell cycle regulation. TFR1 also controls important signaling pathways such as PD1, IL17, IL18, NF-kβ, FOXM1, FOCAL, and JAK-STAT pathways. Besides, TFR1 knockdown in neural stem cells caused epilepsy symptoms in mice, with changes in synaptic GluA2 expression and neurotransmitter release [18-23].
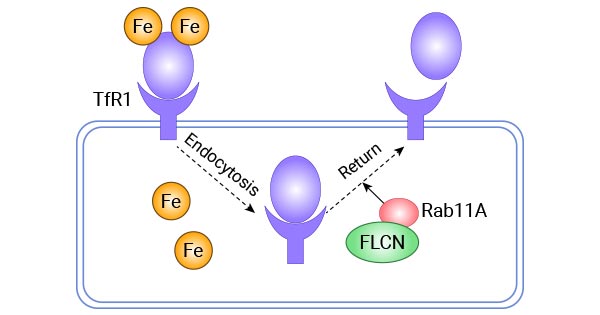
Figure 3. FLCN regulates the recycling and transport of Tf-TFR1 protein via Rab11A [17]
As a regulator of Fe transportation, TFR1 wields substantial influence over numerous health conditions. Specifically, TFRC plays a significant role in the development of anemia, neurodegenerative disorders, and a variety of cancers.
Numerous studies have shown that TFR1 is abnormally expressed in various malignant tumors, including thyroid cancer [24], esophageal squamous cell carcinoma [25], breast cancer [26], liver cancer [27], colon cancer [28], leukemia [29], lung cancer [30], pancreatic cancer [31], and nasopharyngeal cancer [31]. However, there are certain malignancies, such as prostate cancer and testicular cancer, where the expression of TFR1 is not well-defined.
In hepatocellular carcinoma, the expression of TFR1 is correlated with the concentration of methemoglobin and serum thrombospondin [27]. In breast cancer, inhibiting IRP2 expression reduces TFR1 expression and increases ferritin heavy chain expression, which represses the growth of breast cancer cells [26, 32].
In colon cancer, high expression of TFR1 activates the IL-6/IL-11-Stat3 signaling pathway, which promotes proliferation and apoptosis of colonic epithelial cells. This exacerbates colonic mucosa damage and accelerates the progression of colon cancer [28, 33].
Iron metabolism disturbances are known to be one of the pathophysiological mechanisms that trigger neurodegenerative diseases. Iron accumulation in the brain is associated with conditions such as Alzheimer's disease (AD), Parkinson's disease, and amyotrophic lateral sclerosis [34].
AD is one of the most common neurodegenerative diseases characterized by the accumulation of amyloid plaques and loss of certain neurons. Studies have shown that inhibiting the iron uptake proteins TFR1 and TF in the temporal cortex of Alzheimer's model mice, as well as reducing DMT1 expression, can effectively alleviate the iron overload state [35-36].
TFR1 binding to the transferrin (Tf) complex is crucial for cells to acquire iron during erythropoiesis. When there is a shortage of iron or an increase in erythropoiesis, TFR1 expression is increased in response [7, 37].
Clinical studies have confirmed that both expression levels of soluble transferrin receptor (sTFR1) and Tf are significantly higher than normal in thalassemic mice. Further studies have revealed that TFR1 is abnormally highly expressed in β-thalassemia precursor cells. Reducing TFR1 expression effectively regulates ineffective erythropoiesis, improving anemia and alleviating iron overload in affected mice [7, 37].
Studies suggest that TFR1 may also act as a viral receptor and participate in Hepatitis C Virus (HCV) cell entry by mediating HCV-host cell membrane fusion. Given that HCV is a major pathogen causing chronic hepatitis and primary liver cancer, the potential of TFR1 as an HCV antiviral target is subject to interest [38-40]. Furthermore, TFR1 has been discovered to regulate the transport of mGlu1 in neurons and is thought to participate in the mGlu1 signaling pathway, ultimately impacting motor coordination within the cerebellum [41-42].
Currently, several clinical drugs targeting transferrin receptor 1 (TFR1) are under development, including antibody nucleic acid coupled drugs, bisomers, monomers, antibody fusion proteins, ADC drugs, etc.; the indications involve non-small-cell lung cancer, esophageal cancer, Mucopolysaccharidoses Storage Disease (MSD) type, muscular dystrophy, Alzheimer's Disease, and Duchenne Muscular Dystrophy. Among them, JCR Pharmaceuticals Co., Ltd.'sPabinafusp Alfa, an antibody fusion protein targeting IDS and TfR1, has been launched in Japan in 2021. TFR1-based targeted therapeutic strategies have been evolving in recent years. Some studies have utilized TFR1 to improve antibody transport across the blood-brain barrier and to combine with anti-β-amyloid peptide monoclonal antibodies to form specific complex antibodies to improve the therapeutic efficacy in patients with Alzheimer's disease [43-44].
Meanwhile, the anti-TFRC antibody JST-TFR09 and the anti-TFRC monoclonal antibody A24 inhibited iron uptake by tumor cells and induced apoptosis of malignant cells in T-lineage cell leukemia, respectively [45-46]. These studies suggest that TFR1 could be an effective target molecule involved in the clinical treatment of a variety of diseases. In the future, with the deepening and improvement of TFR1-related studies, it is expected to provide patients with more precise and effective treatments, bringing them more benefits and new hopes.
● CUSABIO TFR1 recombinant protein
Recombinant Human Transferrin receptor protein 1(TFRC),partial (Active) (Code: CSB-MP3648HU)
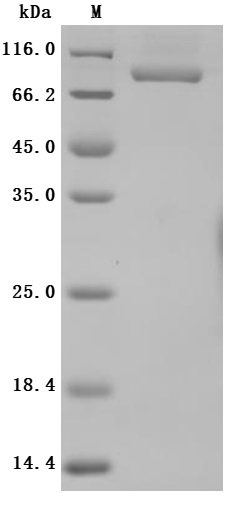
High specificity was validated by SDS-PAGE. SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
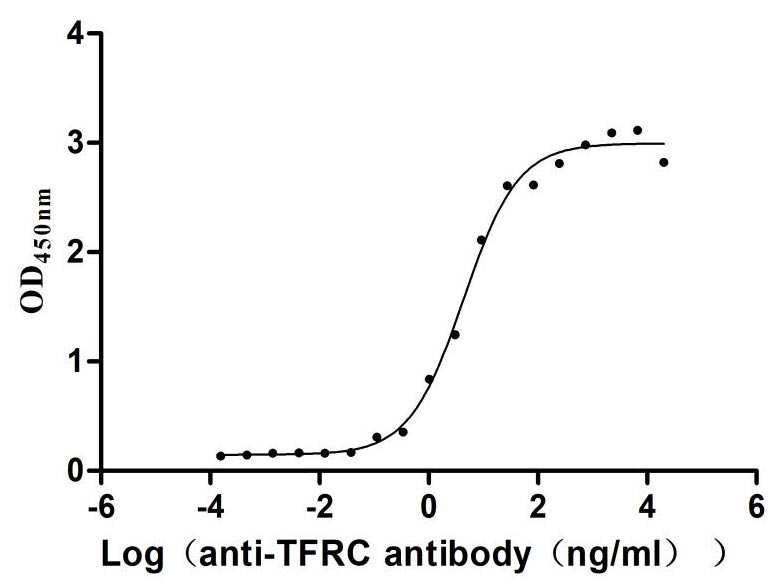
Immobilized Human TFRC at 2μg/mL can bind Anti-TFRC recombinant antibody (CSB-RA023441MA1HU), the EC50 is 3.305-8.220 ng/mL.
● CUSABIO TFR1 antibody
| Product Name | Species Reactivity | Application | Code |
|---|---|---|---|
| TFRC Recombinant Monoclonal Antibody | Human | ELISA | CSB-RA023441MA1HU |
| TFRC Antibody | Human | ELISA, IHC, IF | CSB-PA07219A0Rb |
| TFRC Antibody | Human, Mouse | ELISA, WB, IHC | CSB-PA001477 |
| TFRC Antibody | Human | ELISA, WB | CSB-PA005607 |
| Phospho-TFRC (S24) Antibody | Human, Mouse | ELISA, WB, IHC | CSB-PA010092 |
| TFRC Antibody | Human, Mouse, Rat | ELISA, WB, IHC | CSB-PA023441GA01HU |
| TFRC Antibody | Human, Mouse, Rat | ELISA, WB, IHC | CSB-PA104885 |
| TFRC Antibody | Human, Mouse, Rat | ELISA, WB, IHC | CSB-PA834425 |
● TFR1 ELISA Kit
| Product Name | Sample Types | Detect Range | Code |
|---|---|---|---|
| Mouse transferrin receptor,TFR ELISA Kit | serum, plasma, tissue homogenates | 0.625 ng/mL-40 ng/mL | CSB-E08389m |
| Human soluble transferrin receptor,sTfR ELISA Kit | serum, plasma | Request Information | CSB-E09100h |
References
[1] Transferrin receptor targeting chimeras for membrane protein degradation. Nature, 2024.
[2] Candelaria, Pierre V., et al. "Antibodies targeting the transferrin receptor 1 (TfR1) as direct anti-cancer agents." Frontiers in immunology 12 (2021): 607692.
[3] Jabara, Haifa H., et al. "A missense mutation in TFRC, encoding transferrin receptor 1, causes combined immunodeficiency." Nature genetics 48.1 (2016): 74-78.
[4] Greene, Christopher J., et al. "Transferrin receptor 1 upregulation in primary tumor and downregulation in benign kidney is associated with progression and mortality in renal cell carcinoma patients." Oncotarget 8.63 (2017): 107052.
[5] Kawabata, Hiroshi. "Transferrin and transferrin receptors update." Free Radical Biology and Medicine 133 (2019): 46-54.
[6] Kleven, Mark D., Shall Jue, and Caroline A. Enns. "Transferrin receptors TfR1 and TfR2 bind transferrin through differing mechanisms." Biochemistry 57.9 (2018): 1552-1559.
[7] Li, Huihui, et al. "Decreasing TfR1 expression reverses anemia and hepcidin suppression in β-thalassemic mice." Blood, The Journal of the American Society of Hematology 129.11 (2017): 1514-1526.
[8] Magro, Gaetano, et al. "Aberrant expression of TfR1/CD71 in thyroid carcinomas identifies a novel potential diagnostic marker and therapeutic target." Thyroid 21.3 (2011): 267-277.
[9] Silvestri, Laura, et al. "The extrahepatic role of TFR2 in iron homeostasis." Frontiers in pharmacology 5 (2014): 93.
[10] Tang, Li-Jing, et al. "Ubiquitin-specific protease 7 promotes ferroptosis via activation of the p53/TfR1 pathway in the rat hearts after ischemia/reperfusion." Free Radical Biology and Medicine 162 (2021): 339-352.
[11] Kawabata, Hiroshi. "Transferrin and transferrin receptors update." Free Radical Biology and Medicine 133 (2019): 46-54.
[12] Nadadur, S. S., K. Srirama, and Anuradha Mudipalli. "Iron transport & homeostasis mechanisms: their role in health & disease." Indian Journal of Medical Research 128.4 (2008): 533-544.
[13] Gammella, Elena, et al. "The transferrin receptor: the cellular iron gate." Metallomics 9.10 (2017): 1367-1375.
[14] Tsiftsoglou, Asterios S., Ioannis S. Vizirianakis, and John Strouboulis. "Erythropoiesis: model systems, molecular regulators, and developmental programs." IUBMB life 61.8 (2009): 800-830.
[15] Bayeva, Marina. Novel Regulators of Mitochondrial and Cellular Iron Homeostasis. Diss. Northwestern University, 2012.
[16] Zhao, Lingling, et al. "FLCN is a novel Rab11A-interacting protein that is involved in the Rab11A-mediated recycling transport." Journal of Cell Science 131.24 (2018): jcs218792.
[17] Wang, Xiaojuan, et al. "FLCN regulates transferrin receptor 1 transport and iron homeostasis." Journal of Biological Chemistry 296 (2021).
[18] Ge, Xiaogang, et al. "Treatment with paraquat affects the expression of ferroptosis-related genes." Human & Experimental Toxicology 42 (2023): 09603271231167585.
[19] Wu, Hongrong, et al. "Identification and validation of transferrin receptor protein 1 for predicting prognosis and immune infiltration in lower grade glioma." Frontiers in Molecular Neuroscience 15 (2022).
[20] Wu, Hongrong, et al. "Identification and validation of transferrin receptor protein 1 for predicting prognosis and immune infiltration in lower grade glioma." Frontiers in Molecular Neuroscience 15 (2022): 972308.
[21] Klüssendorf, Malte, et al. "The Golgi-associated PDZ domain protein Gopc/PIST is required for synaptic targeting of mGluR5." Molecular Neurobiology 58.11 (2021): 5618-5634.
[22] Zhou, Jia-Huan, et al. "Ablation of TFR1 in Purkinje cells inhibits mGlu1 trafficking and impairs motor coordination, but not autistic-like behaviors." Journal of Neuroscience 37.47 (2017): 11335-11352.
[23] Warming, Hannah Kate. Haemoglobin neurotoxicity, haptoglobin scavenging and synaptic function in subarachnoid haemorrhage. Diss. University of Southampton, 2023.
[24] Parenti, Rosalba, Lucia Salvatorelli, and Gaetano Magro. "Anaplastic thyroid carcinoma: current treatments and potential new therapeutic options with emphasis on TfR1/CD71." International journal of endocrinology 2014 (2014).
[25] Ye, Jiecheng, et al. "A novel iron (II) phenanthroline complex exhibits anticancer activity against TFR1-overexpressing esophageal squamous cell carcinoma cells through ROS accumulation and DNA damage." Biochemical Pharmacology 166 (2019): 93-107.
[26] Corte-Rodriguez, Mario, et al. "Quantitative analysis of transferrin receptor 1 (TfR1) in individual breast cancer cells by means of labeled antibodies and elemental (ICP-MS) detection." Analytical chemistry 91.24 (2019): 15532-15538.
[27] Xiao, Chong, et al. "Transferrin receptor regulates malignancies and the stemness of hepatocellular carcinoma-derived cancer stem-like cells by affecting iron accumulation." PLoS One 15.12 (2020): e0243812.
[28] Cui, Can, et al. "Downregulation of TfR1 promotes progression of colorectal cancer via the JAK/STAT pathway." Cancer Management and Research 11 (2019): 6323.
[29] Liu, Qian, et al. "Significance of CD71 expression by flow cytometry in diagnosis of acute leukemia." Leukemia & lymphoma 55.4 (2014): 892-898.
[30] Jeong, Seung Min, Sunsook Hwang, and Rho Hyun Seong. "Transferrin receptor regulates pancreatic cancer growth by modulating mitochondrial respiration and ROS generation." Biochemical and biophysical research communications 471.3 (2016): 373-379.
[31] Martínez;nez, Laura E., et al. "Targeting TfR1 with the ch128. 1/IgG1 Antibody Inhibits EBV-driven Lymphomagenesis in Immunosuppressed Mice Bearing EBV+ Human Primary B-cells." Molecular cancer therapeutics 20.9 (2021): 1592-1602.
[32] Chen, Chunli, et al. "Deferoxamine-induced high expression of TfR1 and DMT1 enhanced iron uptake in triple-negative breast cancer cells by activating IL-6/PI3K/AKT pathway." OncoTargets and therapy 12 (2019): 4359.
[33] Huang, Luji, et al. "Iron metabolism in colorectal cancer." Frontiers in Oncology 13 (2023).
[34] Whitnall, Megan, and Des R. Richardson. "Iron: a new target for pharmacological intervention in neurodegenerative diseases." Seminars in pediatric neurology. Vol. 13. No. 3. WB Saunders, 2006.
[35] Yu, Xiaojun, et al. "Decreased iron levels in the temporal cortex in postmortem human brains with Parkinson disease." Neurology 80.5 (2013): 492-495.
[36] Lu, Li-Na, et al. "Expression of iron transporters and pathological hallmarks of Parkinson’s and Alzheimer’s diseases in the brain of young, adult, and aged rats." Molecular neurobiology 54 (2017): 5213-5224.
[37] Cabrera, C., et al. "Relationship between iron deficiency and expression of genes involved in iron metabolism in human myocardium and skeletal muscle." International journal of cardiology 379 (2023): 82-88.
[38] Fillebeen, Carine, and Kostas Pantopoulos. "Hepatitis C virus infection causes iron deficiency in Huh7. 5.1 cells." PLoS One 8.12 (2013): e83307.
[39] Lindenbach, Brett D., and Charles M. Rice. "The ins and outs of hepatitis C virus entry and assembly." Nature Reviews Microbiology 11.10 (2013): 688-700.
[40] Bonkovsky, Herbert L., et al. "Iron and HFE or TfR1 mutations as comorbid factors for development and progression of chronic hepatitis C." Journal of Hepatology 37.6 (2002): 848-854.
[41] Kalinowska, Magdalena. Metabotropic regulation of dendritic spine structural plasticity. Diss. Yeshiva University, 2015.
[42] Zhou, Jia-Huan, et al. "Ablation of TFR1 in Purkinje cells inhibits mGlu1 trafficking and impairs motor coordination, but not autistic-like behaviors." Journal of Neuroscience 37.47 (2017): 11335-11352.
[43] Bray, Natasha. "Transferrin'bispecific antibodies across the blood–brain barrier." Nature Reviews Drug Discovery 14.1 (2015): 14-15.
[44] Pardridge, William M. "Blood–brain barrier drug delivery of IgG fusion proteins with a transferrin receptor monoclonal antibody." Expert opinion on drug delivery 12.2 (2015): 207-222.
[45] Shimosaki, Shunsuke, et al. "Development of a complete human IgG monoclonal antibody to transferrin receptor 1 targeted for adult T-cell leukemia/lymphoma." Biochemical and biophysical research communications 485.1 (2017): 144-151.
[46] Candelaria, Pierre V., et al. "Antibodies targeting the transferrin receptor 1 (TfR1) as direct anti-cancer agents." Frontiers in immunology 12 (2021): 607692.


Comments
Leave a Comment