Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
Lately, the journal Seminars in Immunology published an intriguing article titled "IL-1R8: A Molecular Brake for Anti-Tumor and Anti-Viral Activity of NK Cells and ILCs" [1]. The study highlights the significance of IL-1R8, also known as SIGIRR, as a crucial regulatory molecule controlling the immune response in tumors and viruses. Notably, SIGIRR is a newly discovered member of the Toll-IL-1R (TIR) superfamily, which includes both the Interleukin-1 receptors (ILRs) and Toll-like receptors (TLRs).
In addition, SIGIRR negatively regulates the signaling pathways of ILRs and TLRs, effectively controlling inflammation and immune responses in a variety of diseases including infections, tumors, and autoimmune disorders. As a result, SIGIRR holds promise as a key player in the research of immune response and inflammation mechanisms. Moreover, its potential as a novel anti-inflammatory 'brake' makes it an attractive target for clinical drug research!
1. What is the Toll-IL-1R (TIR) Superfamily?
3. How does SIGIRR Negatively Regulate the TLRs and IL-1Rs Signaling Pathway?
Toll-IL-1R (TIR) is a new group of signaling receptors. Toll-like receptors (TLRs) and Interleukin-1 receptors (ILRs) are important immune receptors that share a conserved sequence called Toll/IL-1R (TIR) structural domain in the cytoplasmic region. Therefore, they are classified into the Toll-IL-1R (TIR) family. The human TIR superfamily can be divided into three groups: the Toll-like receptor family, the IL-1R family, and My D88 (Figure 1) [2-3].
The TLRs subfamily consists of various receptors, namely TLR1, TLR2, TLR3, TLR4, TLR5, TLR6, TLR7, TLR8, and TLR9 etc. The ILRs subfamily includes several receptors, such as IL-1R1, IL-18R1, T1/ST2, IL-1Rrp2, and certain cytokines like IL-1, IL-18, and IL-33. Additionally, there are other IL-1 family members, including AcP (IL-1RAcP, IL-18RAcP, and IL-1RAcPb), IL-1F6, IL-1F8, and IL-1F9. Studies have shown that TIR family members are crucial for natural immunity against foreign pathogens. However, there exist orphan receptors in the TIR family whose functions and ligands remain unknown, like SIGIRR [1-3].
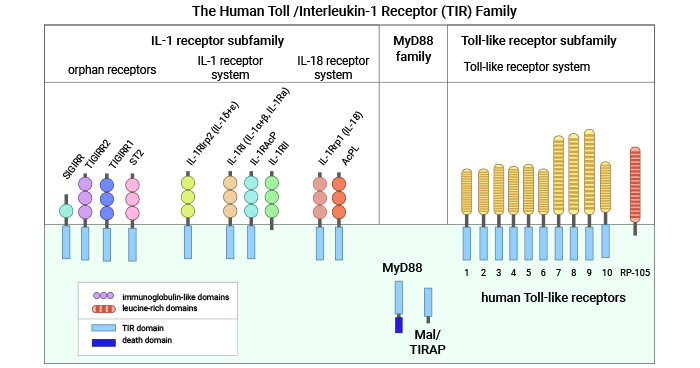
Figure 1. Human Toll-IL-1R (TIR) superfamily [3]
Single Ig IL-1-related receptor (SIGIRR), also named Interleukin-1 receptor8 (IL-1R8), is a member of the TIR superfamily and was first discovered by Thomassen et al. in 1999. SIGIRR gene is located on chromosome 11, and SIGIRR protein comprises a single Ig extracellular structural domain (aa17-112), a transmembrane structural domain (aa117-139), an intracellular TIR conserved structural domain (aa166-305), and an intracellular tail region that measures 95aa in length (Figure 2) [4-7].
SIGIRR is structurally similar to IL-1R and TLRs, but the structural specificity of SIGIRR is demonstrated by the fact that SIGIRR has only one Ig extracellular structural domain, while IL-1R has three; the intracellular TIR structural domain of SIGIRR lacks the amino acids Ser447 and Tyr536, which are necessary for signaling. Given the unique molecular structure of SIGIRR, SIGIRR cannot activate downstream signaling pathways in the inflammatory response [4-7].
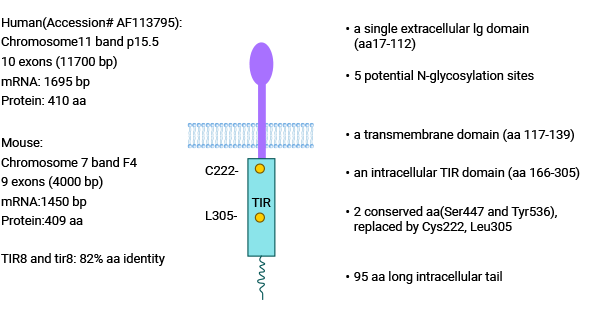
Figure 2. SIGIRR Structure [4]
SIGIRR expression is found in various tissues, including the lung, liver, parathyroid gland, gastrointestinal tract, and kidney. It plays a crucial role in regulating inflammation. Inflammatory states are characterized by reduced SIGIRR expression levels, leading to increased p38 activation and decreased expression of USP13. When stimulated with endotoxin LPS, USP13 expression decreases, resulting in an increase in SIGIRR ubiquitination and decreased stability. The widespread expression of SIGIRR in different tissues and organs enables its negative regulatory mechanism in inflammation [8-10].
SIGIRR, an orphan receptor, has its ligand not fully elucidated; however, some studies suggest that IL-1F5 is its ligand in glial cells. SIGIRR has the capability to inhibit the signaling of T cell differentiation associated with IL-1 family members and TLRs-mediated activation, resulting in a crucial regulatory role in inflammation, tumor-associated inflammation, and autoimmunity [11-13].
Evidence from mouse studies under different conditions robustly demonstrates the negative regulatory role of SIGIRR. Moreover, emerging clinical data support its relevance to human disease. In a nutshell, the regulatory role of SIGIRR in inflammatory immune responses holds great clinical significance [11-13].
SIGIRR is a unique protein that negatively regulates TLRs and IL-1Rs in inflammation-related diseases, affecting downstream targets such as IL-1R1, IL-1R5/IL-18Rα, IL-1R4/ST2, TLR4, TLR7, TLR9, TLR3, and TLR1/2. It inhibits TIR structural domain-induced NF-κB and JNK activation, but the mechanism is not yet fully understood [14-16].
SIGIRR mainly inhibits IL-1-induced signaling, Pam3Cys-induced TLR2 signaling pathway, endotoxin (LPS) with CpG DNA-induced TLR4 and TLR9, and activation of TLR3 and TLR1/2 signaling pathways. It achieves this through two mechanisms: interfering with IL-1RI and IL-1RAcP to form a dimer and blocking TRAM dimerization to interfere with TRIF signaling [17-19].
Three-dimensional modeling shows that the TIR structural domain competes with the MyD88 dimer and inhibits signal transduction, particularly its BB-loop. SIGIRR also inhibits MyD88 by blocking TRAM dimerization to interfere with TRIF signaling. This affects TLR3 signaling as well as TLR4-TRAM and TRIF-TRAM interactions. Additionally, SIGIRR regulates JNK and mTOR phosphorylation in Th17 cells and NK cells and IL-1 or TLR agonist-driven mTOR phosphorylation in intestinal epithelial cells (Figure 3) [20-22].
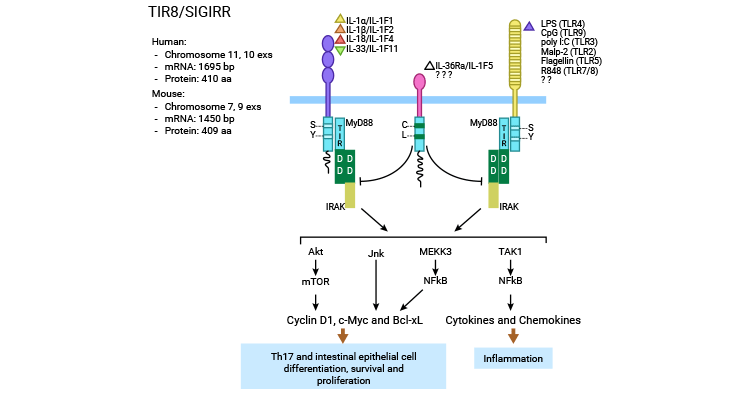
Figure 3. SIGIRR negatively regulate the TLRs and IL-1Rs signaling pathway [22]
SIGIRR has a key role in regulating inflammatory immune-related diseases. It helps to prevent tissue damage, promotes recovery from infectious diseases, reduces risk and severity of autoimmune diseases, and inhibits tumor development. But, it may also assist in the spread of tumor cells.
Abnormal SIGIRR expression is linked to severe inflammatory damage in body tissues. In Mycobacterium tuberculosis infections, mice with a defective SIGIRR gene experience excessive inflammation and increased mortality due to deregulation of its inhibitory effects on IL-1 and TNF-α [23-24].
In other infection models, such as acute lung infections caused by Pseudomonas aeruginosa, Candida albicans, or Aspergillus fumigatus, and urinary tract infections caused by Escherichia coli, low SIGIRR expression or silencing leads to heightened organismal susceptibility, excessive release of pro-inflammatory cytokines, and increased mortality [13].
Furthermore, SIGIRR-deficient mice exhibit more severe β-amyloid peptide-induced synaptic and cognitive dysfunction. However, up-regulation of SIGIRR alleviates Abeta-mediated impairment of hippocampal synaptic activity over a long-term potentiation (LTP) [17, 25].
ILRs and TLRs play a part in the pathophysiology of autoimmune diseases and allergic inflammation, with IL-1 being a key factor in promoting Th17 differentiation and activation in multiple autoimmune diseases. On the other hand, SIGIRR acts as an inhibitor to regulate IL-1-dependent Th17 differentiation, expansion, and effector functions, as well as various IL-1-dependent signaling pathways in T cells, including the mTOR pathway associated with Th17 proliferation [21, 25].
Lack of SIGIRR is associated with an increased risk of autoimmune disease and positively correlates with disease severity. In terms of allergic inflammation, the role of SIGIRR is still debated. In IL-33-dependent airway allergic inflammation, mice without SIGIRR exhibit pulmonary inflammation, splenomegaly, elevated serum IL-5 and IL-13 levels, and increased production of Th2 cytokines [26-27].
Inflammatory environments can promote tumor growth, and the relationship between tumors and inflammation is affected by genetic susceptibility genes for endogenous malignancies and exogenous inflammation or infection. These mechanisms are linked to the activation of NF-κB, STAT3, and HIF-1α.
SIGIRR is a factor that suppresses tumor-associated inflammatory responses, and its deletion affects the development and progression of inflammation-associated tumors. In a colon cancer model, SIGIRR-deficient mice exhibit increased activation of NF-κB signaling in their colon tissue. This promotes tumor formation by affecting target genes critical for cell proliferation and survival, such as Cyclin D1 and Bcl-xL [28-29]. Additionally, various cytokines (e.g., TNF-α, IL-6, and IFN-γ) and chemokines (e.g., MIP-2, MCP-1, and CXCL1/KC) were detected in the colonic tissue of these mice, indicating an altered immune response during tumor development [28-29].
SIGIRR inhibits the IL-18 pathway, which affects natural killer (NK) cell differentiation and functional maturation. The inhibitory effect of SIGIRR on tumor development has been demonstrated in both a diethylnitrosamine-induced liver cancer model and a mouse breast cancer model (Figure 4) [1, 30-32].
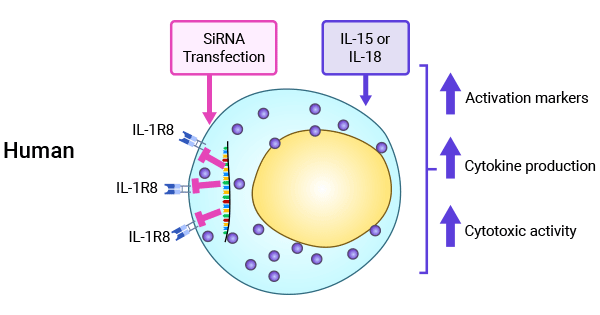
Figure 4. The importance of SIGIRR (IL-1R8) in regulating NK cell function [4]
Recent advancements highlight SIGIRR's exceptional role as a negative regulator in TIR family-mediated inflammatory signaling pathways. Its clinical potential has been demonstrated in various immune-related diseases, and over-activation can cause inflammation and disrupt immune homeostasis. SIGIRR's negative regulation in inflammatory immune response maintains the balance of downstream signaling pathways mediated by the TIR superfamily. Despite the positive contribution of TIR members to immune response-related diseases, their regulatory balance is not well-understood. Therefore, SIGIRR boasts great significant clinical value as a "brake" molecule that regulate inflammation and immune response!
To fully support researchers and pharmaceutical companies in their research on SIGIRR in immune-mediated inflammatory diseases, CUSABIO presents SIGIRR protein to support your research on the mechanism of SIGIRR or its potential clinical value (click for the full list of SIGIRR products: SIGIRR Proteins; SIGIRR antibodies).
Recombinant Human Single Ig IL-1-related receptor(SIGIRR),partial (Code: CSB-MP743558HU)
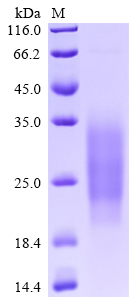
High purity was validated by SDS-PAGE. (Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
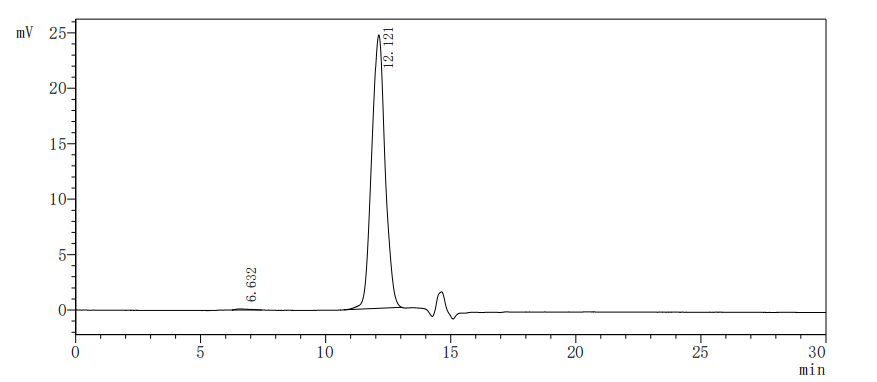
The purity of Human SIGIRR was greater than 95% as determined by SEC-HPLC.
References
[1] Mariotti F R, Supino D, Landolina N, et al. IL-1R8: A molecular brake of anti-tumor and anti-viral activity of NK cells and ILC[C]//Seminars in Immunology Academic Press, 2023, 66: 101712.
[2] Carty M, Kearney J, Shanahan K A, et al. Cell survival and cytokine release after inflammasome activation is regulated by the Toll-IL-1R protein SARM[J]. Immunity, 2019, 50(6): 1412-1424. e6.
[3] Martin M U, Wesche H. Summary and comparison of the signaling mechanisms of the Toll/interleukin-1 receptor family[J]. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research, 2002, 1592(3): 265-280.
[4] Huang Xun. Experimental study of SIGIRR regulating HMGB1-induced inflammatory response of alveolar epithelial cells A549. Diss. Fourth Military Medical University, 2012.
[5] Thomassen E, Renshaw B R, Sims J E. Identification and characterization of SIGIRR, a molecule representing a novel subtype of the IL-1R superfamily[J]. Cytokine, 1999, 11(6): 389-399.
[6] Lech M, Skuginna V, Kulkarni O P, et al. Lack of SIGIRR/TIR8 aggravates hydrocarbon oil-induced lupus nephritis[J]. The Journal of Pathology: A Journal of the Pathological Society of Great Britain and Ireland, 2010, 220(5): 596-607.
[7] Drexler S K, Kong P, Inglis J, et al. SIGIRR/TIR-8 is an inhibitor of toll-like receptor signaling in primary human cells and regulates inflammation in models of rheumatoid arthritis[J]. Arthritis & Rheumatism, 2010, 62(8): 2249-2261.
[8] Li L, Wei J, Li S, et al. The deubiquitinase USP13 stabilizes the anti-inflammatory receptor IL-1R8/Sigirr to suppress lung inflammation[J]. EBioMedicine, 2019, 45: 553-562.
[9] Li L, Wei J, Suber T L, et al. IL-37-induced activation of glycogen synthase kinase 3β promotes IL-1R8/Sigirr phosphorylation, internalization, and degradation in lung epithelial cells[J]. Journal of cellular physiology, 2021, 236(8): 5676-5685.
[10] Wang Q, Sun Z, Xia W, et al. Role of USP13 in physiology and diseases[J]. Frontiers in Molecular Biosciences, 2022, 9: 977122.
[11] Garlanda C, Anders H J, Mantovani A. TIR8/SIGIRR: an IL-1R/TLR family member with regulatory functions in inflammation and T cell polarization[J]. Trends in immunology, 2009, 30(9): 439-446.
[12] Bertilaccio M T S, Simonetti G, Dagklis A, et al. Lack of TIR8/SIGIRR triggers progression of chronic lymphocytic leukemia in mouse models[J]. Blood, The Journal of the American Society of Hematology, 2011, 118(3): 660-669.
[13] Riva F, Bonavita E, Barbati E, et al. TIR8/SIGIRR is an interleukin-1 receptor/toll like receptor family member with regulatory functions in inflammation and immunity[J]. Frontiers in immunology, 2012, 3: 322.
[14] Heinig K, Sperandio M. Interleukin-1R8: balancing inflammation and hemostasis in platelets[J]. Cardiovascular Research, 2016, 111(4): 307-309.
[15] Molgora M, Barajon I, Mantovani A, et al. Regulatory role of IL-1R8 in immunity and disease[J]. Frontiers in immunology, 2016, 7: 149.
[16] Giannoudaki E, Stefanska A M, Lawler H, et al. SIGIRR negatively regulates IL-36-driven psoriasiform inflammation and neutrophil infiltration in the skin[J]. The Journal of Immunology, 2021, 207(2): 651-660.
[17] Costello D A, Carney D G, Lynch M A. α-TLR2 antibody attenuates the Aβ-mediated inflammatory response in microglia through enhanced expression of SIGIRR [J]. Brain, behavior, and immunity, 2015, 46: 70-79.
[18] Qin J, Qian Y, Yao J, et al. SIGIRR inhibits interleukin-1 receptor- and toll-like receptor 4-mediated signaling through different mechanisms[J]. Journal of Biological Chemistry, 2005, 280(26): 25233-25241.
[19] Zhang C, Wu X, Zhao Y, et al. SIGIRR inhibits toll-like receptor 4, 5, 9-mediated immune responses in human airway epithelial cells[J]. Molecular biology reports, 2011, 38: 601-609.
[20] Zhao R, Song C, Liu L, et al. Single immunoglobulin and Toll-interleukin-1 receptor domain containing molecule protects against severe acute pancreatitis in vitro by negatively regulating the Toll-like receptor-4 signaling pathway: A clinical and experimental study[J]. Molecular Medicine Reports, 2020, 22(4): 2851-2859.
[21] Gulen M F, Kang Z, Bulek K, et al. The receptor SIGIRR suppresses Th17 cell proliferation via inhibition of the interleukin-1 receptor pathway and mTOR kinase activation[J]. Immunity, 2010, 32(1): 54-66.
[22] Jiang Keguo. The regulation of SIGIRR on the activation of NF-κB in human renal tubular epithelial cells, the feedback regulation of NF-κB on SIGIRR and the preliminary study on the mechanism of SIGIRR regulation of EMT. Diss. Anhui Medical University, 2015.
[23] Horne D J, Randhawa A K, Chau T T H, et al. Common polymorphisms in the PKP3-SIGIRR-TMEM16J gene region are associated with susceptibility to tuberculosis [J]. Journal of Infectious Diseases, 2012, 205(4): 586-594.
[24] Garlanda C, Di Liberto D, Vecchi A, et al. Damping excessive inflammation and tissue damage in Mycobacterium tuberculosis infection by Toll IL-1 receptor 8/single Ig IL-1-related receptor, a negative regulator of IL-1/TLR signaling[J]. The Journal of Immunology, 2007, 179(5): 3119-3125.
[25] Costello D A, Watson M B, Cowley T R, et al. Interleukin-1α and HMGB1 mediate hippocampal dysfunction in SIGIRR-deficient mice[J]. Journal of Neuroscience, 2011, 31(10): 3871-3879.
[26] Martin M U. Special aspects of interleukin-33 and the IL-33 receptor complex[C]//Seminars in immunology. Academic Press, 2013, 25(6): 449-457.
[27] Lunding L, Webering S, Vock C, et al. IL-37 requires IL-18Rα and SIGIRR/IL-1R8 to diminish allergic airway inflammation in mice[J]. Allergy, 2015, 70(4): 366-373.
[28] Xiao H, Gulen M F, Qin J, et al. The Toll-interleukin-1 receptor member SIGIRR regulates colonic epithelial homeostasis, inflammation and tumorigenesis[J]. Immunity, 2007, 26(4): 461-475.
[29] Liu J, Chen Y, Liu D, et al. Ectopic expression of SIGIRR in the colon ameliorates colitis in mice by downregulating TLR4/NF-κB overactivation[J]. Immunology letters, 2017, 183: 52-61.
[30] Che Y Y, Shi X, Zhong X D, et al. Resveratrol prevents liver damage in MCD-induced steatohepatitis mice by promoting SIGIRR gene transcription[J]. The Journal of Nutritional Biochemistry, 2020, 82: 108400.
[31] Molgora M, Bonavita E, Ponzetta A, et al. IL-1R8 is a checkpoint in NK cells regulating anti-tumour and anti-viral activity[J]. Nature, 2017, 551(7678): 110-114.
[32] Bodaszewska-Lubas M, Liao Y, Zegar A, et al. Dominant-Negative Form of SIGIRR: SIGIRRΔE8 Promotes Tumor Growth Through Regulation of Metabolic Pathways[J]. Journal of Interferon & Cytokine Research, 2022, 42(9): 482-492.


Comments
Leave a Comment