Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
A research paper entitled “HSF1 promotes CD69+Treg differentiation to inhibit colitis progression” was recently published in the journal Theranostics [1]. The study found that knockdown of CD69 induced autoimmune diseases such as inflammatory bowel disease. In addition, HSF1 promotes CD69+Tregs differentiation by activating the CD69 transcription, which is critical for the immunosuppressive function of Tregs. In particular, for patients undergoing treatment for autoimmune diseases, increasing the number of CD69+Treg cells is expected to alleviate disease symptoms.
CD69 is considered as a pivotal regulator that plays an important role in the immune system as the earliest molecule expressed for lymphocyte activation. An increasing number of studies have shown that CD69 functions as a key regulatory in hematologic diseases and autoimmune disorders. As for the biological functions of CD69 and its mechanism of action in immune regulation, further research and exploration are needed. Let's explore together!
CD69, also known as AIM, EA-1, Leu23, and MLR-3, is a type II transmembrane glycoprotein expressed on the surface of lymphocytes upon activation. Human CD69 consists of 199 amino acids [2], which includes a 40-residue intracellular region, a 21-residue transmembrane region, and a 138-residue extracellular region containing C-type lectins with Ca2+-dependent carbohydrate-recognition domain (CRD), making it a member of the C-type lectin receptor family. The terminal Ser/Thr in the intracellular compartment is phosphorylated, and CD69 is associated with CD23 and CD72 [3-4].
CD69 is also part of the NK cell-specific genetic factor complex. However, unlike NKR-P1, Ly-49, and NKG2 molecules that are exclusively expressed on NK cells, CD69 can be expressed on various cells, indicating its diverse properties (Figure 1) [2].
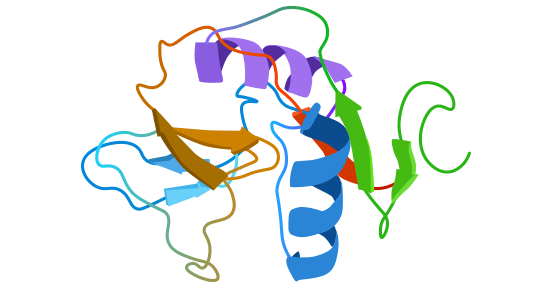
Figure 1. Schematic diagram of CD69 structure [2]
CD69 is a surface antigen that is expressed by lymphocytes upon activation. Further research has revealed that it is also expressed on nearly all blood cells following induction. CD69 can be induced in a variety of cell types, including T cells, B cells, NK cells, monocytes, neutrophils, eosinophils, and thymocytes, etc. The fact that CD69 is only expressed upon cell activation makes it an ideal marker molecule for identifying activated cells [5-7].
In the case of T cell activation, CD69 is one of several representative molecules, including IL-2 receptor CD25 and transferrin receptor CD71. In addition to its role as a marker for cell activation, CD69 can also function as a co-stimulatory signal. Studies have shown that CD69+T cells accompanied by IL-12β, IL-18α, and IL-16β mRNA levels increase, and in the presence of recombinant IL-12 or IL-18, promotes interferon-γ (IFN-γ) production. However, recent research has revealed that while CD69 promotes cell activation, it may also be closely associated with apoptosis [8-9].
Although the specific ligand of CD69 remains unclear, some studies suggest that CD69 may be involved in Galectin-1 (Gal-1) binding, which regulates Th17 cells (Figure 2) [8]. In summary, CD69 cross-linking generates intracellular signaling and prompts multiple immune responses, with a wide range of effects on the biological functions of hematopoietic cells.
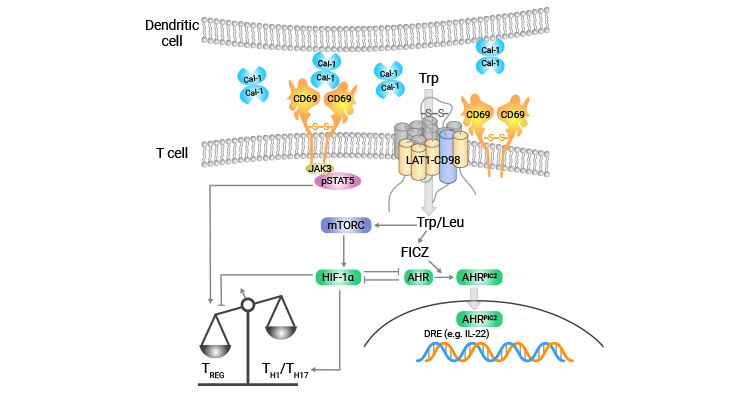
Figure 2. CD69 may bind to Galectin-1 (Gal-1) to regulate Th17 cells [8]
CD69 is a multifunctional immunomodulatory molecule that plays crucial roles in the activation and differentiation of various hematopoietic cells. Despite its discovery, the precise mechanism underlying CD69-mediated cellular activation and apoptosis remains poorly understood. Initial investigations have primarily concentrated on the role of CD69 in cell activation.
Resting T cells do not express CD69, but when exposed to stimulation by anti-CD3/TCR (T cell receptor) and activators of CD2, CD28, and protein kinase C (PKC) such as PMA and PHA, T cells can be induced to upregulate CD69 expression.
The presence of the CD3/TCR complex is essential for T cell activation initiated by CD69 (Figure 3) [10]. In addition, PMA participates in intracellular signaling pathways by activating GTP ligation and Ras, which are crucial for inducing CD69 expression on T cells.
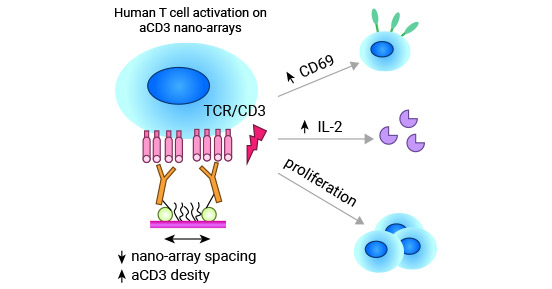
Figure 3. CD3/TCR complex is necessary in CD69-mediated T-cell activation [10]
NK cells also do not express CD69 in the resting state. However, activation of NK cells is induced by cross-linking CD16 and treatment with IL-2, IL-7, IL-12, IFNA, or PMA, which leads to the expression of CD69 [10-11]. This process involves various protein kinases, including multiple protein kinase C (PKC) and protein tyrosine kinase (PTK), that participate in diverse signaling pathways [10-11]. The cytotoxic activity of NK cells initiated by CD69 is modulated by CD94 [12]. Besides, it has been reported that CD69 activates extracellular signal-regulated kinase ERK. However, CD69-initiated ERK activation is negatively regulated by the inhibitory receptor CD94/NKG2-A [13].
For B cells in spleen and lymph nodes, lipopolysaccharide (LPS) induces molecular expression of CD69. Conversely, in peripheral blood, CD69 expression in eosinophil EOS can be induced through exposure to cytokines such as IL-2, IL-3, IL-5, IL-13, GM-CSF, and IFN-γ in vitro [14-15].
CD69 is closely associated with apoptosis in vivo. T cells that exhibit high expression of CD69 upon activation in vivo are more susceptible to rapid spontaneous apoptosis in vitro. Notably, CD69-mediated apoptosis differs from CD95-mediated apoptosis in its specific characteristics. While CD95-mediated apoptosis affects almost all cell types, CD69-mediated apoptosis primarily occurs in activated T cells, eosinophils EOS, and monocytes.
Studies indicate that CD69-mediated monocyte apoptosis is linked to three distinct signaling pathways: 1) NO-dependent pathway; 2) pathway associated with the phospholipase A2/phospholipid oxidase cascade reaction; 3) pathway influenced by PTX-sensitive G proteins; 4) CD69-mediated TGF-β synthesis has also been identified as a contributing factor [16].
Caspases play a crucial role in apoptosis. In EOL-1 human eosinophilic cell line, CD45, CD45R, CD45RB, CD95, and CD69 monoclonal antibody resulted in increased apoptosis. This effect was inhibited by caspase-8 and caspase-9 inhibitors, while CD69 cross-linking led to activation of caspase-3 and caspase-8. Treatment with anti-CD69 monoclonal antibodies induced apoptosis in EOS and reduced bcl-2 expression. However, in the presence of GM-CSF, the intracellular concentration of bcl-2 remained unchanged. Notably, CD69-induced apoptosis decreased bcl-2 expression in GM-CSF cultured CD69+ cells, suggesting that CD69-induced apoptosis is associated with bcl-2 [17].
The mechanism of apoptosis by mature T cell activation may be FAS. In vivo activation of tonsillar T cells with high CD69 expression has been shown to increase their susceptibility to spontaneous and rapid apoptosis. Additionally, several cellular factors including CXCL12/SDF-1, TIL16, IL17, IL12, and IL15 have been found to inhibit apoptosis. Conversely, inhibition of phosphatidylinositol 3 kinase (PI3K) has been shown to accelerate apoptosis, suggesting that multiple exogenous signals contribute to an endogenous increase in apoptosis susceptibility of activated CD69+ T cells in vivo [18].
CD69 is a critical component of the T cell-mediated immune response. Its expression promotes T cell activation by increasing intracellular Ca2+ concentration, upregulating the expression of cytokines such as TNF-α, IFN-γ, IL-2, while also acting as a co-stimulatory signal. CD69 plays a vital role in the activation, proliferation, and differentiation of various blood cells and is involved in the pathogenesis of hematological diseases and immune-related disorders, according to available studies.
A significant increase in the expression of activation marker molecules such as CD69 and CD71 on CD4+ and CD8+ T cells was found in a subpopulation of peripheral blood lymphocytes from a variety of hematologic malignancies, including lymphoma, acute or chronic leukemia, and multiple myeloma, indicating the presence of T cell activation in such diseases.
Acute myeloid leukemia (AML) is characterized by the abnormal proliferation and differentiation of hematopoietic stem cells, leading to the presence of immature leukemic cells that rapidly displace normal hematopoietic tissue. This results in various symptoms such as anemia, infection, and bleeding. One of the key factors contributing to this disease is the reduced expression of CD69, a protein involved in cytokine secretion.
As a result of decreased CD69 expression, the production of cytokines like IL2 and TNF-α is also diminished, which impairs the formation of effector T cells and can compromise the immune response. However, studies have shown that following complete remission with chemotherapy, CD69 expression rates dramatically increase, indicating that AML patients have suppressed T-cell activation and impaired immune function [19].
Chronic Lymphocytic Leukemia (CLL) is a type of lymphocytic leukemia that involves the malignant clonal proliferation of CD5+ CD19+ B lymphocytes. In vitro stimulation of B cells with mitogens in CLL patients resulted in similar expression of CD69 compared to normal groups. But CD95 expression was impaired. Further investigations have shown that CLL patients express surface activating antigens like CD23, CD25, CD69, and CD71, with CD69 expression levels being closely associated with peripheral blood lymphocyte counts and worse clinical staging in CLL patients [20].
In myelodysplastic syndromes (MDS) monocytes, CD69 expression is significantly increased, which may be associated with immune dysfunction in MDS patients. High CD69 expression may lead to decreased clearance of pathogens by monocytes, thus making MDS patients more susceptible to infection.
The majority of research on CD69 has centered around autoimmune diseases such as systemic lupus erythematosus and rheumatoid arthritis. These studies have demonstrated that CD69 is highly expressed in the peripheral blood of individuals with autoimmune diseases, and following effective treatment, CD69 expression levels significantly decrease. Additionally, there is a notable correlation between disease progress and CD69 levels.
Systemic Lupus Erythematosus (SLE) is an autoimmune disease characterized by dysregulated T and B lymphocyte function. In stimulation experiments, lymphocytes from SLE patients displayed increased CD69 expression in response to various stimuli compared to cells from healthy individuals. Additionally, lymphocytes from SLE patients exhibited moderate proliferation upon exposure to a range of stimuli. Notably, no significant differences in CD69 expression were observed in SLE patients with CD4+ and CD8+ T cells, suggesting that CD69 is involved in the activation of both T cell subsets and likely functions to transmit early-stage activation signals [22].
In rheumatoid arthritis (RA) patients' synovial fluid, CD69 expression was found to be elevated in both CD4+ and CD8+ T lymphocytes, indicating activation of both T cell subsets. Analysis of CD69 expression in the synovial fluid of RA, seronegative spondyloarthropathy (SSd), and crystalline arthritis (CAA) patients revealed no significant differences in the percentage of CD69+ lymphocytes among the three groups. However, the highest level of CD69 expression was observed in RA patients' synovial fluid. Furthermore, lymphocyte CD69 expression in RA patients was found to be positively correlated with synovial fluid IL-15 levels [23-24].
In mice with autoimmune myocarditis, CD69 was found to play a negative regulatory role in myocardial inflammation by modulating cardiac-specific Th17 cells. This subset of immune cells is involved in the development of several inflammatory diseases, including myocarditis. In vivo, CD69 was observed to regulate T cell differentiation to Th17 cells through Jak3/Stat5 pathway [25].
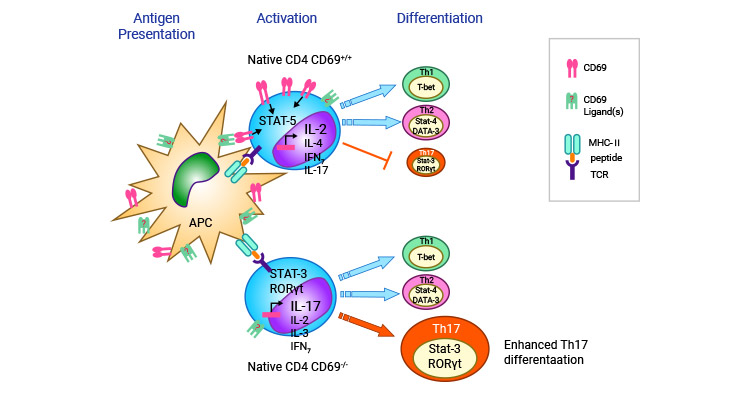
Figure 4. CD69 regulates the differentiation of T cells to Th17 cells [25]
Research has demonstrated that patients with coronary heart disease (CHD) exhibit high expression of CD69, DKK-1, IL-10, IL-6, TC, TG, and LDL, which have been observed to increase in proportion to the severity of coronary stenosis lesions [26-27]. Previous studies have also indicated that T cells in the airways of asthma patients express CD69, which induces apoptosis of these cells by regulating TGF-β production and secretion upon activation [28]. The activation of apoptotic cells through CD69 provides new insight into potential treatments for asthma.
Additionally, other researchers have observed that the expression levels of CD69 in the peripheral blood of patients with solid tumors (such as gastric cancer, liver cancer, and colorectal cancer) were significantly reduced and negatively correlated with tumor stage, but rebounded after surgical resection of the lesion or chemotherapy [29-31].
GeneFrontier Corp. is developing a monoclonal antibody drug (GFC-101) that targets CD69 and shows promise for treating conditions such as irritable bowel syndrome, rheumatoid arthritis, and respiratory disorders. CD69 as an early activated protein can trigger immune responses. Other drugs or antibodies targeting CD69 expression are being investigated, including in the treatment of CLL, like blocking B cell receptors to lower CD69 levels may be a way to reduce the number of patients who do not respond well to Bendamustine [32]. Recent studies have revealed potential ligands (such as Gla-1 and S1PR1) of CD69 [8, 33]. As further research is conducted on CD69, it is anticipated that innovative immunotherapeutic therapies aimed at CD69 will emerge, with potential applications in the treatment of hematological and autoimmune disorders.
To fully support researchers and pharmaceutical companies in their research on CD69 in hematological and autoimmune disorders, CUSABIO presents CD69 active proteins ( CSB-MP004952HU and CSB-EP004952HU-B) to support your research on the mechanism of CD69 or its potential clinical value (click for the full list of CD69 products: CD69 Proteins; CD69 antibodies).
CD69 proteins:
Recombinant Human Early activation antigen CD69(CD69),partial (Active)
Recombinant Human Early activation antigen CD69(Cd69),partial,Biotinylated
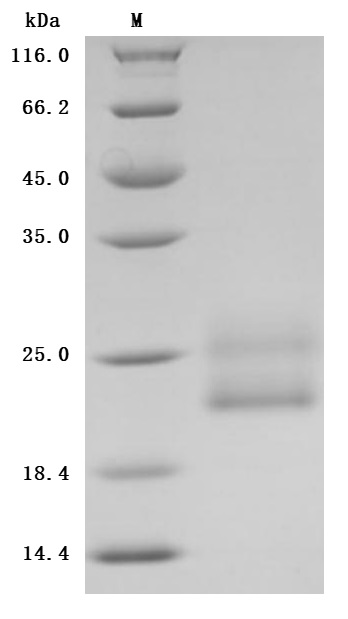
The purity was greater than 96.7% as determined by SDS-PAGE. (Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
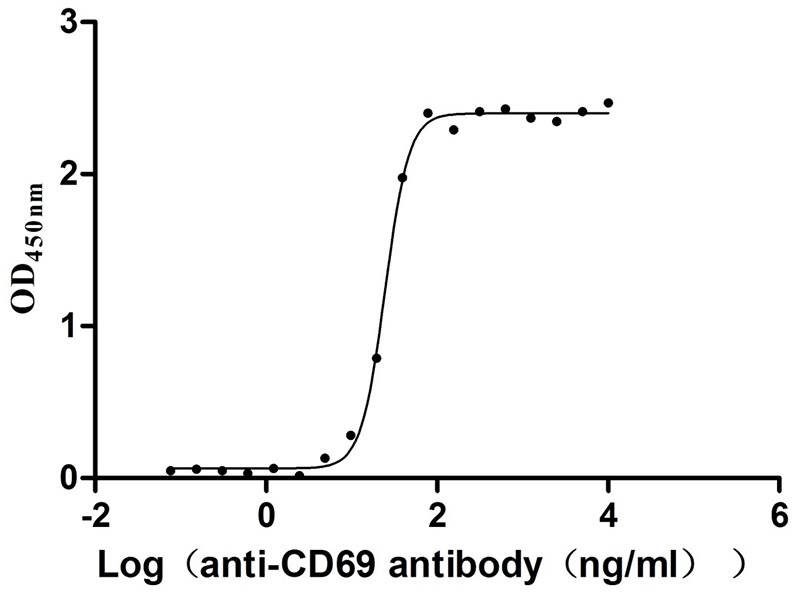
Immobilized Human CD69 at 2μg/mL can bind Anti-CD69 recombinant antibody (CSB-RA004952MA1HU), the EC50 is 23.17-26.04 ng/mL
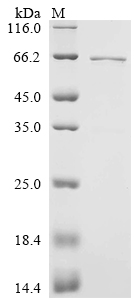
The purity was greater than 90% as determined by SDS-PAGE.(Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
CD69 Stable Transfer Cell Line:
References
[1] Yu, Lei, et al. "HSF1 promotes CD69+ Treg differentiation to inhibit colitis progression." Theranostics 13.6 (2023): 1892.
[2] Dohnálek, Jan, and Tereza Skálová. "C-type lectin-(like) fold-protein-protein interaction patterns and utilization." Biotechnology Advances 58 (2022): 107944.
[3] Koyama-Nasu, Ryo, et al. "The cellular and molecular basis of CD69 function in anti-tumor immunity." International Immunology 34.11 (2022): 555-561.
[4] Vandeveer, George H., et al. "Discovery of structurally diverse reversible BTK inhibitors utilized to develop a novel in vivo CD69 and CD86 PK/PD mouse model." Bioorganic & Medicinal Chemistry Letters 80 (2023): 129108.
[5] Lauzurica, Pilar, et al. "Phenotypic and functional characteristics of hematopoietic cell lineages in CD69-deficient mice." Blood, The Journal of the American Society of Hematology 95.7 (2000): 2312-2320.
[6] Chen, Chun-Kai, et al. "Increased expressions of CD69 and HLA-DR but not of CD25 or CD71 on endometrial T lymphocytes of nonpregnant women." Human immunology 42.3 (1995): 227-232.
[7] Peggs, Karl S., et al. "Immunotherapy with CD25/CD71-allodepleted T cells to improve T-cell reconstitution after matched unrelated donor hematopoietic stem cell transplant: a randomized trial." Cytotherapy 25.1 (2023): 82-93.
[8] Cibrián, Danay, and Francisco Sánchez-Madrid. "CD69: from activation marker to metabolic gatekeeper. "European journal of immunology 47.6 (2017): 946-953.
[9] Agaugué, Sophie, et al. "Human natural killer cells exposed to IL-2, IL-12, IL-18, or IL-4 differently modulate priming of naive T cells by monocyte- derived dendritic cells." Blood, The Journal of the American Society of Hematology 112.5 (2008): 1776-1783.
[10] Matic, J., Deeg, J., Scheffold, A., Goldstein, I., & Spatz, J. P. (2013). Fine tuning and efficient T cell activation with stimulatory aCD3 nanoarrays. nano letters, 13(11), 5090-5097.
[11] Tian, Yunfei, et al. "Development of a Monoclonal Antibody to Pig CD69 Reveals Early Activation of T Cells in Pig after PRRSV and ASFV Infection." Viruses 14.6 (2022): 1343.
[12] Borrego, F., et al. "CD69 is a stimulatory receptor for natural killer cell and its cytotoxic effect is blocked by CD94 inhibitory receptor." Immunology 97.1 (1999): 159.
[13] Zingoni, Alessandra, et al. "CD69-triggered ERK activation and functions are negatively regulated by CD94/NKG2-A inhibitory receptor." European journal of immunology 30.2 (2000): 644-651.
[14] Ishizaki, Shunsuke, et al. "Role of CD69 in acute lung injury." life sciences 90.17-18 (2012): 657-665.
[15] Lamana, Amalia, et al. "CD69 modulates sphingosine-1-phosphate-induced migration of skin dendritic cells." Journal of Investigative Dermatology 131.7 (2011): 1503-1512.
[16] Ramı́rez, Rafael, et al. "CD69-induced monocyte apoptosis involves multiple nonredundant signaling pathways. "Cellular immunology 172.2 (1996). 192-199.
[17] Aouad, Salah M., et al. "Caspase-3 is a component of Fas death-inducing signaling complex in lipid rafts and its activity is required for complete caspase-8 activation during Fas-mediated cell death." The Journal of Immunology 172.4 (2004): 2316-2323. -8 activation during Fas-mediated cell death." The Journal of Immunology 172.4 (2004): 2316-2323.
[18] Deng, Caishu, Elzbieta Goluszko, and Premkumar Christadoss. "Fas/Fas ligand pathway, apoptosis, and clonal anergy involved in systemic acetylcholine receptor T cell epitope tolerance." The Journal of Immunology 166.5 (2001): 3458-3467.
[19] Qiu, Guo, Xi Xu, and Qifa Liu. "CD69 Marks Leukemic Regenerating Cells and Regulates Their Metabolism in AML." Blood 140.Supplement 1 (2022): 8780-8782.
[20] Del Poeta, Giovanni, et al. "CD69 is independently prognostic in chronic lymphocytic leukemia: a comprehensive clinical and biological profiling study." Haematologica 97.2 (2012): 279-287.
[21] Davison, Glenda M., Nicolas Novitzky, and Rygana Abdulla. "Monocyte derived dendritic cells have reduced expression of co-stimulatory molecules but are able to stimulate autologous T-cells in patients with MDS." Hematology/oncology and stem cell therapy 6.2 (2013): 49-57.
[22] Portales-Perez, D., et al. "Abnormalities in CD69 expression, cytosolic pH and Ca2+ during activation of lymphocytes from patients with systemic lupus erythematosus." Lupus 6.1 (1997): 48-56.
[23] Atzeni, Fabiola, et al. "CD69 expression on neutrophils from patients with rheumatoid arthritis." clinical and experimental rheumatology 22.3 (2004) : 331-334.
[24] Afeltra, A., et al. "Expression of CD69 antigen on synovial fluid T cells in patients with rheumatoid arthritis and other chronic synovitis." Annals of the rheumatic diseases 52.6 (1993): 457-460.
[25] Martín, Pilar, et al. "CD69 association with Jak3/Stat5 proteins regulates Th17 cell differentiation." Molecular and cellular biology 30.20 (2010). 4877-4889.
[26] Ivanovska, Nina, and Petya Dimitrova. "Bone resorption and remodeling in murine collagenase-induced osteoarthritis after administration of glucosamine." Arthritis research & therapy 13.2 (2011): 1-13.
[27] Peng, Jianqiao, and Yi Xiang. "Value analysis of CD69 combined with EGR1 in the diagnosis of coronary heart disease. "Experimental and Therapeutic Medicine 17.3 (2019): 2047-2052.
[28] Martín, Pilar, et al. "The leukocyte activation antigen CD69 limits allergic asthma and skin contact hypersensitivity." Journal of Allergy and Clinical Immunology 126.2 (2010): 355-365.
[29] Cai, Yang, Wei Zhu, and Guanghua Feng. "Alteration of CD69 and HLA-DR-positive T Cells in Patients with Gastric Cancer after Operations and its Clinical Significances." Journal of Medical Research (2006).
[30] Bruni, Elena, et al. "Intrahepatic CD69+ Vδ1 T cells re-circulate in the blood of patients with metastatic colorectal cancer and limit tumor progression ." Journal for Immunotherapy of Cancer 10.7 (2022).
[31] Tang, Kaihua, et al. "CD69 serves as a potential diagnostic and prognostic biomarker for hepatocellular carcinoma." Scientific Reports 13.1 (2023). 7452.
[32] Montraveta, Arnau, et al. "CD69 expression potentially predicts response to bendamustine and its modulation by ibrutinib or idelalisib enhances cytotoxic effect in chronic lymphocytic leukemia." Oncotarget, 2015, vol. 7, num. 5, p. 5507-5520 (2015).
[33] Chen, Hongwen, et al. "Transmembrane protein CD69 acts as an S1PR1 agonist." Elife 12 (2023): e88204.


Comments
Leave a Comment