
Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
| Code | CSB-MP023969HU |
| Abbreviation | Recombinant Human TNFRSF11B protein (Active) |
| MSDS | |
| Size | $9.9 |
| Promotion |

|
| Order now | |
| Image |
|
| Have Questions? | Leave a Message or Start an on-line Chat |

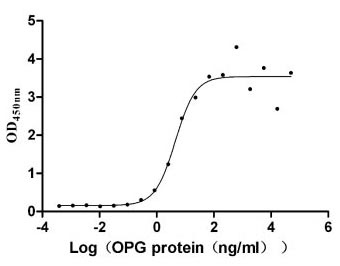
TNFRSF11B (osteoprotegerin) serves as a soluble decoy receptor for RANKL, making it a critical regulator of osteoclastogenesis and a target of growing interest in cancer-associated bone remodeling research. This recombinant human TNFRSF11B covers the full-length mature protein (aa 22–401) and carries a C-terminal hFc1-Flag tag, produced in mammalian cells to preserve native folding and proper disulfide bond formation essential for accurate ligand engagement. Functional ELISA confirms robust RANKL-binding activity with an EC50 of 2.651–7.646 ng/ml against immobilized TNFSF11, providing a reliable positive control for receptor-ligand interaction assays and supporting use in competitive inhibition studies, blocking antibody screening, and affinity characterization by SPR or BLI. Purity exceeding 90% by SDS-PAGE combined with endotoxin levels below 1.0 EU/μg satisfies the quality thresholds commonly required for drug discovery workflows targeting small-molecule or biologic inhibitors of the RANKL–OPG axis.
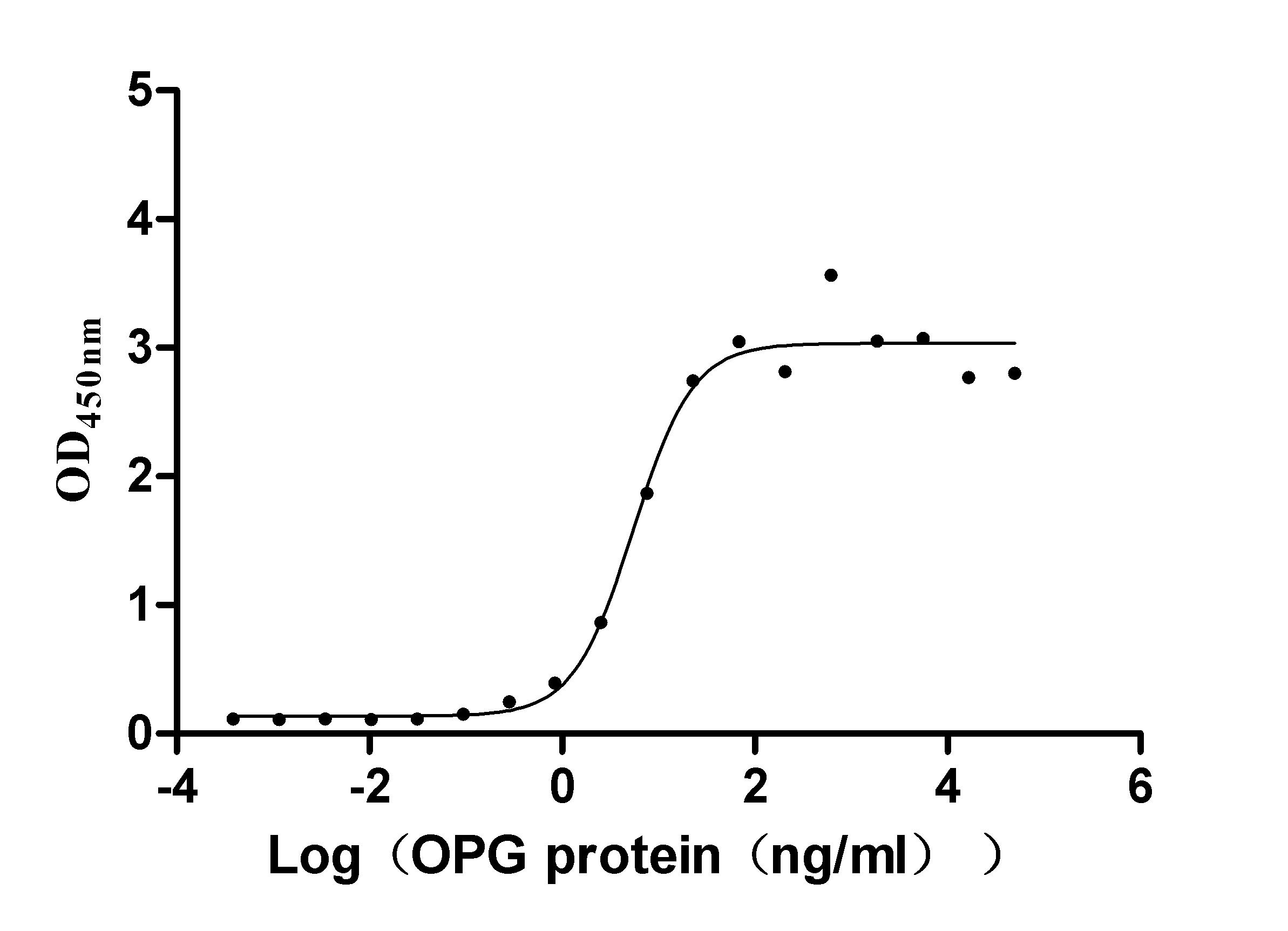
Applications : Binding assay/Protein-protein interaction
Review: After receiving the product, functional ELISA experiments were conducted with Human TNFSF11, and the EC50 was 3.935-7.640 ng/ml, indicating good binding activity. Subsequently used for antibody screening, ideal clones were screened, and the stability between batches was good!
By Anonymous

