Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
Cytokines are molecules produced by immune cells that play a crucial role in regulating the immune response. Some cytokines are pro-inflammatory while others are anti-inflammatory. Numerous inflammation-related cytokines are emerging as novel targets for assessing susceptibility to tumors and immune disorders. Importantly, the interleukin IL-6 family assumes a vital pro-inflammatory role, emerging as a significant player in influencing the inflammatory response.
Recent years have witnessed an intensified exploration of the IL-6 family, shedding light on OSM. As a result, OSM's significance in tumor development and immunomodulation has garnered escalating attention. Ample evidence emphasizes the promising potential of OSM as a robust target for upcoming diagnostics and precision therapies in various diseases, including tumors, bone disorders, cardiovascular issues, autoimmune diseases, and more.
1. Interleukin-6 Family Cytokines
3. What are the Receptors for OSM?
4. How Does OSM Binding to Its Receptors Regulate the JAK/STAT3 and MAPK Signaling Pathways?
6. The Clinical Potential of OSM Targeted Therapy
7. CUSABIO OSM Recombinant Proteins & Antibodies for Research Use
The interleukin-6 (IL-6) family is a group of cytokines that can be produced by a wide range of cells. The IL-6 family of cytokines, including IL-6, IL-11, IL-27, LIF, OSM, CNTF, CT-1, and CLCF1, is produced by various cells, functioning in autocrine, paracrine, and endocrine signaling. These cytokines play crucial roles in regulating metabolism, inflammation, and immune balance. Research indicates that they exhibit dual effects on tumors—both promoting and inhibiting tumorigenesis and progression. Recently, the IL-6 cytokine family has gained significant attention in tumor immunology research [1-2].
Oncostatin-M (OSM) is an IL-6 family cytokine. First identified in 1986 from U937 histiocytic lymphoma cells, it inhibits melanoma cell growth. The OSM gene resides on chromosome 22q12, comprising three exons and two introns adjacent to LIF. OSM's protein holds 239 to 263 amino acids, folding into a four-helix bundle akin to other IL-6 family proteins (Figure 1) [3]. The human OSM polypeptide has 252 amino acids, with a 25-residue N-terminus signal peptide and 32-residue C-terminus. STAT5 response element plays a pivotal role in OSM transcription. Cytokines like IL-2, IL-3, EPO, GM-CSF, among others, activate STAT5, boosting OSM expression. Furthermore, prostaglandin E2/PG-E2 can upregulate OSM transcription [3-5].
OSM is secreted mainly by activated monocytes, macrophages, T lymphocytes, and dendritic cells. It holds a crucial role in maintaining the hematopoietic microenvironment, regulating G-CSF and SDF-1 to support progenitor cell function. In addition, OSM influences CNS development, liver regeneration, cardiomyocyte behavior, cytokine production by endothelial cells, and angiogenesis. Current in vitro studies reveal OSM's broad impact on hematopoiesis, cell dynamics, inflammation, metabolism, tumorigenesis, and immune regulation [3-6].
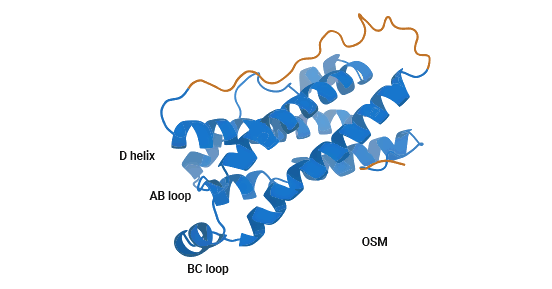
Figure 1. OSM Structure [3]
OSM shares a gp130 receptor subunit with various other cytokines within the IL-6 superfamily. However, its affinity for gp130 is notably low. Receptors for these IL-6 cytokines inherently include the gp130 glycoprotein as a signaling receptor subunit. Although gp130 is expressed across cell types, it alone is not capable of cytokine binding. Receptor expression for distinct IL-6 cytokine family members varies by cell type [7-8].
OSM engages two distinct receptor complexes, distinguishing itself from other IL-6 family cytokines. In humans, two types of functional OSM receptor are known: type I and type II. Type I OSM receptors (gp130/LIFRβ) constitute a heterodimer featuring a gp130 molecule alongside a LIF receptor subunit. This heterodimeric configuration binds both LIF and gp130, initiating signaling. In contrast, type II OSM receptors (gp130/OSMRβ) comprise a gp130 molecule paired with an OSM receptor β subunit. Type II receptors are exclusive to OSM and fail to bind LIF and other family cytokines (Figure 2) [8].
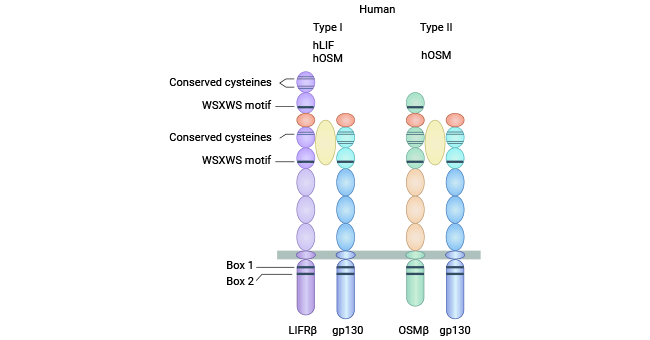
Figure 2. type I OSM receptor (gp130/LIFRβ) and type II OSM receptor (gp130/OSMRβ) [8]
OSM triggers JAK/STAT3 and MAPK signaling pathways by binding to distinct receptor pairs (gp130/LIFRβ; gp130/OSMRβ). These pathways are pivotal in governing a spectrum of biological responses (Figure 3) [9].
The JAK/STAT3 pathway initiates when OSM binds to the gp130/OSM receptor beta subunit complex. This triggers receptor dimerization, mutual phosphorylation, and Janus kinase (JAK) activation. Activated JAK then phosphorylates gp130's cytoplasmic tyrosine residues. Signal transduction factors and transcription activators (STAT) recognize these phosphorylated receptors, leading to STAT autophosphorylation. Phosphorylated STAT molecules form dimers, entering the nucleus as active transcription factors that modulate specific gene expression [9].
In parallel, gp130's tyrosine residue phosphorylation attracts tyrosine phosphatases and facilitates binding to growth factor receptor-binding protein 2 (Grb2). This enables Grb2 to attach to SOS protein, activating SOS. SOS triggers Ras protein's transition from GDP to GTP, activating Ras. Activated Ras activates Raf-1, which in turn phosphorylates MAPK. Activated MAPK then phosphorylates extracellular signal-regulated protein kinase (ERK) 1/2. These phosphorylated ERK1/2 molecules move to the nucleus, targeting specific transcription factors to modulate gene expression and prompt distinct biological outcomes [9].
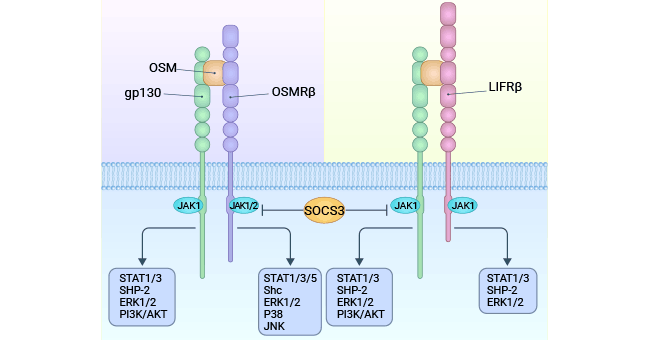
Figure 3. OSM signals through two types of receptors (gp130/LIFRb) and gp130/OSMRb complexes [9]
OSM's functions and mechanisms remain largely unknown. Initially known for inhibiting melanoma growth, OSM's regulatory roles in organ development, tissue recovery, and repair have emerged. However, OSM's effects on cell proliferation and differentiation can differ or even oppose each other. Current research mainly explores OSM's relevance in conditions like tumors, bone metabolism, hematopoiesis, and chronic inflammation.
OSM cytokines play diverse roles in tumors, tied to cancer stage and aggressiveness. In normal cells and early tumors, OSM suppresses proliferation and halts progression. However, in highly malignant or metastatic cases, increased OSM associates with poor outcomes.
Consider hepatocellular carcinoma cells as an example. OSM activates the STAT3 pathway and upregulates c-Myc oncogene expression, inducing cellular senescence. This inhibits hepatocellular carcinoma proliferation (Figure 4) [9]. In advanced tumors, OSM often fuels proliferation. Oncogene-triggered cellular aging curtails abnormal cell growth while preserving tissue health. However, this mechanism relies on proteins like p53 or Rb, frequently impaired in advanced cancer cells. Thus, OSM's dual effects on liver cancer proliferation can be attributed to these varying contexts [9-10].
OSM's influence extends to various tumors like melanoma, lung, breast, ovarian, and gastric cancer [9-13]. In breast cancer, OSM downregulates E-cadherin, upregulating slug and snail expressions. For glioma, OSM regulates a mesenchymal phenotype, boosting migration and invasion, correlating with malignancy and low patient survival [15]. OSM significantly affects TGFβ signaling, promoting migration and invasion of gastroenteropancreatic neuroendocrine tumors (GEP-NENs) [16].
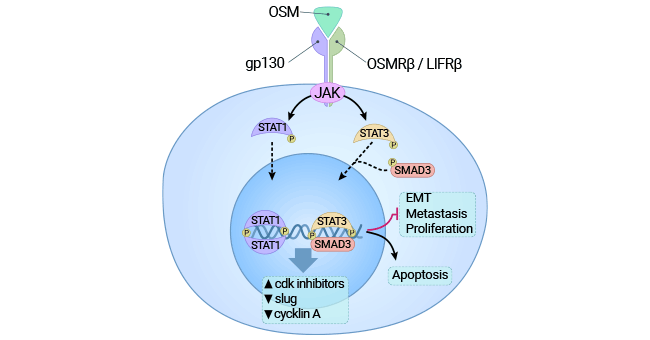
Figure 4. Binding of OSM receptors activates the JAK3/STAT1 pathway and exerts cytotoxic effects on cancer cells [9]
Both LIF and OSM impact rat cranial cell osteoblast differentiation. In mouse osteoblasts, binding with BMP-2 increased alkaline phosphatase, implying OSM recruits BMP during periosteal bone formation. Mouse cranial explants stimulated with OSM released collagen and regulated Osteoprotegerin (OPG), curbing bone resorption. In rheumatoid arthritis, synoviocyte OSM indirectly degraded proteins, triggering joint PGE2-mediated matrix degradation [17-18].
Utilizing OSM/IL-1 or OSM/TNF stimulation models, OSM induced MMP-10 in chondrocytes and synovial fibroblasts. MMP-10, in turn, activated fibroblast activating protein-α in chondrocytes, bolstering cartilage metabolism [18]. Furthermore, an OSM antibody regulated OSM's interaction with IL-1 and TNF in arthritic mice, substantially curbing collagen breakdown and alleviating inflammation. This presents a novel avenue for arthritis treatment [19].
Inflammatory bowel disease (IBD), including Crohn's disease (CD) and ulcerative colitis (UC), involves OSM playing both pro-inflammatory and anti-inflammatory roles in the inflammation response. OSM's levels increase in colonic lesions of active CD patients, aiding epithelial repair [20]. Damaged intestinal mucosa in IBD patients displays heightened OSM and OSMR receptor expression, and OSM triggers the expression of inflammatory factors like IL-6 and ICAM-1 [20].
Active UC patients treated with diverse anti-TNF drugs revealed varying OSM expression levels. The group unresponsive to anti-TNF-α antibody Infliximab exhibited the highest OSM expression [21-22]. Hence, Inhibiting OSM has the potential to decrease the expression of inflammatory factors associated with inflammatory bowel disease (IBD) in the intestinal mucosa. This highlights the potential of targeting OSM as a therapeutic approach.
Some investigations have implied OSM's involvement in cardiovascular disease development. OSM spurs endothelial cells to produce IL-6, G-CSF, GM-CSF, and adhesion molecules, facilitating neutrophil adhesion [23-24]. Moreover, OSM modulates COX-2 to enhance neutrophil adhesion, and it fosters microcirculatory endothelial cell angiogenesis by regulating angiopoietin-2 (ANG-2) [23-24]. Additionally, OSM impacts vascular smooth muscle cells, inducing cell proliferation and the secretion of VEGF, IL-6, and COX-2 [25].
In atherosclerosis, OSM affects endothelial cells, vascular smooth muscle cells, and angiogenesis [26-27]. OSM's role in myocardial infarction and ventricular remodeling is similarly intricate, enhancing cardiac function through diverse mechanisms, yet potentially exhibiting adverse effects in certain instances [26-27].
Furthermore, OSM has been linked to a range of metabolic-immune and neurological disorders [28-29]. For instance, either alone or in conjunction with other GP130 cytokines, OSM can disrupt normal adipose tissue metabolism, inducing lipid metabolism irregularities that may contribute to diabetes [30]. OSM's influence extends to adipocyte differentiation inhibition and lipid metabolism disorders through the Ras/Extracellular Signal-regulated Kinase (Ras/ERK) and STAT5 pathways, by regulating CCAAT enhancer-binding protein β (C/EBP β) [31-32].
OSMR-deficient mice, exhibiting diminished TRPV1/P2X3/OSMRβ neuron expression, also display reduced responsiveness to mechanical and thermal stimuli. This suggests OSM's potential as a neuroprotective cytokine, encouraging exploration of its therapeutic prospects in central nervous system (CNS) disorders [33].
OSM is a cytokine belonging to the interleukin IL-6 family with a range of biological functions. Notably, OSM’s activation occurs uniquely through the OSMRβ/gp130 (type II OSM receptor) pathway, which fuels its distinct impacts. While the mechanics behind its diverse biological actions and interactions with other cytokines remain unclear, this area of research requires further investigation. Despite these uncertainties, present research suggests that OSM and its antagonists hold the potential for promising therapeutic applications in clinical disease treatments. Further studies of the molecular mechanisms and physiological functions of OSM may lead to the development of novel therapeutic strategies for a wide range of diseases.
The OSM protein is a multifunctional cytokine that plays a critical role in physiological mechanisms and disease progression. OSM activates multiple signaling pathways and mediates various biological effects, including cell growth, differentiation, apoptosis, inflammation, and immune response. Although the role of OSM in cancer and autoimmunity is complex and context-dependent, OSM-based therapies have promising therapeutic potential in the treatment of certain cancers and inflammatory diseases.
To fully support researchers and pharmaceutical companies in their research on OSM in autoimmune diseases, tumors, or other diseases, CUSABIO presents OSM active proteins & antibodies to support your research on the mechanism of OSM or its potential clinical value.
CUSABIO Oncostatin-M (OSM) protein
● Recombinant Human OSM, partial (Active) (Code: CSB-MP017260HU1)
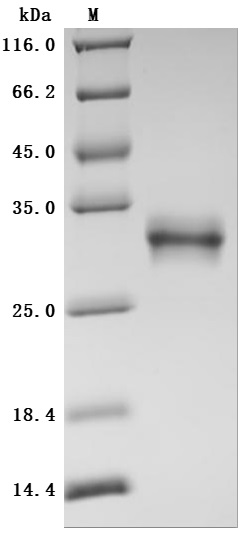
The high purity was greater than 95% as determined by SDS-PAGE.
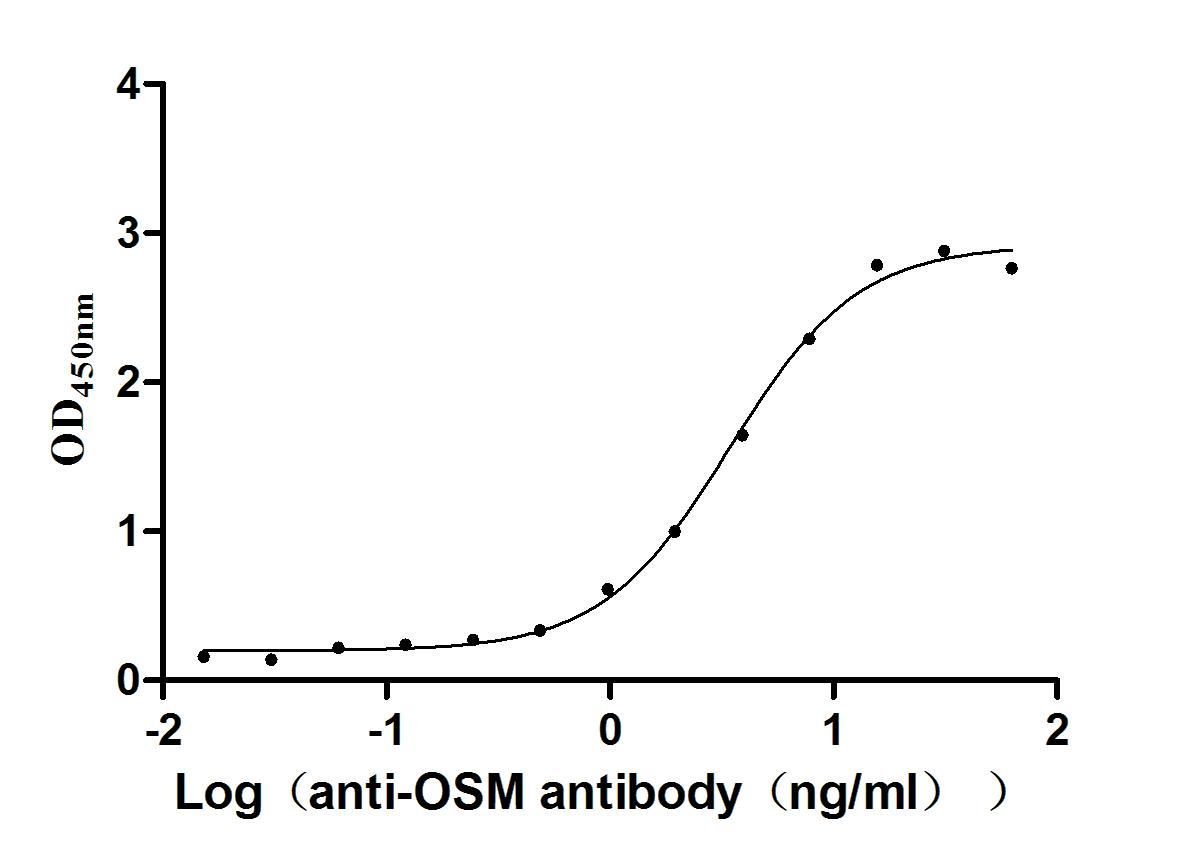
Immobilized Human OSM at 2μg/mL can bind Anti-OSM recombinant antibody (CSB-RA017260MA1HU). The EC50 is 3.048-3.860 ng/mL.
CUSABIO Oncostatin-M (OSM) Antibody
OSM Recombinant Monoclonal Antibody (Code: CSB-RA017260MA1HU)
References
[1] Tanaka, Toshio, Masashi Narazaki, and Tadamitsu Kishimoto. "Interleukin (IL-6) immunotherapy." cold spring harbor perspectives in biology 10.8 ( 2018): a028456.
[2] Jones, Simon A., Jürgen Scheller, and Stefan Rose-John. "Therapeutic strategies for the clinical blockade of IL-6/gp130 signaling." The Journal of clinical investigation 121.9 (2011): 3375-3383.
[3] Du, Qingqing, Yan Qian, and Weiwei Xue. "Molecular simulation of oncostatin M and receptor (OSM-OSMR) interaction as a potential therapeutic target for inflammatory bowel disease." Frontiers in Molecular Biosciences 7 (2020): 29.
[4] Bruce, A. Gregory, Peter S. Linsley, and Timothy M. Rose. "Oncostatin M." Progress in growth factor research 4.2 (1992): 157-170.
[5] Yoshimura, Akihiko, et al. "Mouse oncostatin M: an immediate early gene induced by multiple cytokines through the JAK-STAT5 pathway." The EMBO journal 15.5 (1996): 1055-1063.
[6] Botelho, Fernando M., et al. "Pulmonary expression of oncostatin M (OSM) promotes inducible BALT formation independently of IL-6, despite a role for IL- 6 in OSM-driven pulmonary inflammation." The Journal of Immunology 191.3 (2013): 1453-1464.
[7] Mosley, Bruce, et al. "Dual oncostatin M (OSM) receptors: cloning and characterization of an alternative signaling subunit conferring OSM-specific receptor activation." Journal of Biological Chemistry 271.51 (1996): 32635-32643.
[8] Grant, Susan L., and C. Glenn Begley. "The oncostatin M signalling pathway: reversing the neoplastic phenotype?" Molecular Medicine Today 5.9 (1999): 406-412.
[9] Masjedi, Ali, et al. "Oncostatin M: A mysterious cytokine in cancers." International Immunopharmacology 90 (2021): 107158.
[10] Caligiuri, Alessandra, et al. "Oncostatin M: From intracellular signaling to therapeutic targets in liver cancer." Cancers 14.17 (2022): 4211.
[11] M Stephens, Jacqueline, and Carrie M Elks. "Oncostatin M: potential implications for malignancy and metabolism. "Current pharmaceutical design 23.25 (2017): 3645-3657.
[12] Douglas, Andrea M., et al. "Oncostatin M induces the differentiation of breast cancer cells." International journal of cancer 75.1 (1998): 64-73.
[13] Junk, Damian J., et al. "Oncostatin M promotes cancer cell plasticity through cooperative STAT3-SMAD3 signaling." Oncogene 36.28 (2017): 4001-4013.
[14] West, N. R., J. I. Murray, and P. H. Watson. "Oncostatin-M promotes phenotypic changes associated with mesenchymal and stem cell-like differentiation in breast cancer." Oncogene 33.12 (2014): 1485-1494.
[15] Halfter, Hartmut, et al. "Inhibition of growth and induction of differentiation of glioma cell lines by oncostatin M (OSM)." Growth Factors 15.2 (1998): 135-147.
[16] Xue, Jingwen, et al. "TCF-3-mediated transcription of lncRNA HNF1A-AS1 targeting oncostatin M expression inhibits epithelial-mesenchymal transition via TGFβ signaling in gastroenteropancreatic neuroendocrine neoplasms." Aging (Albany NY) 13.10 (2021): 14065.
[17] Guihard, Pierre, et al. "Induction of osteogenesis in mesenchymal stem cells by activated monocytes/macrophages depends on oncostatin M signaling." Stem cells 30.4 (2012): 762-772.
[18] Hui, Wang, et al. "A model of inflammatory arthritis highlights a role for oncostatin M in pro-inflammatory cytokine-induced bone destruction via RANK/ RANKL." Arthritis Res Ther 7 (2004): 1-8.
[19] Le Goff, Benoit, et al. "Oncostatin M acting via OSMR, augments the actions of IL-1 and TNF in synovial fibroblasts." Cytokine 68.2 (2014): 101-109.
[20] West, Nathaniel R., et al. "Oncostatin M drives intestinal inflammation and predicts response to tumor necrosis factor-neutralizing therapy in patients with inflammatory bowel disease." Nature medicine 23.5 (2017): 579-589.
[21] Ito, Hiroaki. "IL-6 and Crohn's disease." Current Drug Targets-Inflammation & Allergy 2.2 (2003): 125-130.
[22] Verstockt, Sare, Bram Verstockt, and Séverine Vermeire. "Oncostatin M as a new diagnostic, prognostic and therapeutic target in inflammatory bowel disease (IBD)." Expert Opinion on Therapeutic Targets 23.11 (2019): 943-954.
[23] Albasanz-Puig, Adaia, et al. "Oncostatin M is expressed in atherosclerotic lesions: a role for Oncostatin M in the pathogenesis of atherosclerosis." Atherosclerosis 216.2 (2011): 292-298.
[24] Kubin, Thomas, et al. "The role of oncostatin M and its receptor complexes in cardiomyocyte protection, regeneration, and failure." International Journal of Molecular Sciences 23.3 (2022): 1811.
[25] Repovic, Pavle, et al. "Oncostatin-M induction of vascular endothelial growth factor expression in astroglioma cells." Oncogene 22.50 (2003): 8117- 8124.
[26] Gusakova, Anna M., et al. "Association of increased oncostatin M with adverse left ventricular remodeling in patients with myocardial infarction." Journal of Medical Biochemistry 41.4 (2022): 441.
[27] Han, Hui, et al. "Oncostatin M promotes infarct repair and improves cardiac function after myocardial infarction." American journal of translational research 13.10 (2021): 11329.
[28] Richards, Carl D. "The enigmatic cytokine oncostatin m and roles in disease." International Scholarly Research Notices 2013 (2013).
[29] Guo, Sen, et al. "Oncostatin M confers neuroprotection against ischemic stroke." Journal of Neuroscience 35.34 (2015): 12047-12062.
[30] Sanchez-Infantes, David, et al. "Oncostatin m is produced in adipose tissue and is regulated in conditions of obesity and type 2 diabetes. "The Journal of Clinical Endocrinology & Metabolism 99.2 (2014): E217-E225.
[31] Miyaoka, Yuichiro, et al. "Oncostatin M inhibits adipogenesis through the RAS/ERK and STAT5 signaling pathways." Journal of Biological Chemistry 281.49 (2006): 37913-37920.
[32] Zhou, Yue, et al. "Blockage of oncostatin M-induced LDL receptor gene transcription by a dominant-negative mutant of C/EBPβ." Biochemical Journal 397.1 (2006): 101-108.
[33] Morikawa, Yoshihiro. "Oncostatin M in the development of the nervous system." Anatomical science international 80 (2005): 53-59.


Comments
Leave a Comment