Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
| Code | CSB-MP5112MOW |
| Abbreviation | Recombinant Rhesus macaque CCN2 protein (Active) |
| MSDS | |
| Size | $138 |
| Order now | |
| Image |
|
| Have Questions? | Leave a Message or Start an on-line Chat |
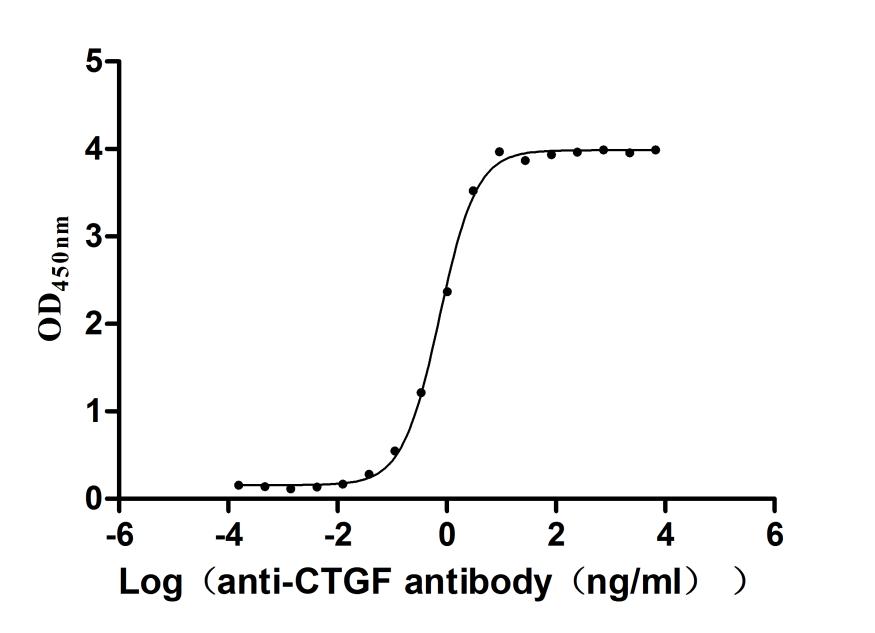
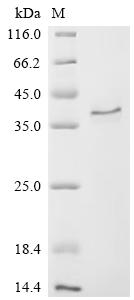
CCN2, also known as connective tissue growth factor, orchestrates fibroblast proliferation, extracellular matrix remodeling, and angiogenesis across wound healing and fibrotic disease models. This full-length mature protein (aa 27–349) demonstrates robust antibody-binding activity with an EC50 of 0.69–0.78 ng/mL in functional ELISA, providing a quantitative benchmark for antibody characterization studies, receptor-ligand competition assays, and ELISA standard curve generation. Mammalian cell expression preserves the native glycosylation and disulfide architecture critical for CCN2's interaction with integrins and heparan sulfate proteoglycans, supporting its use in cell proliferation assays, osteogenic and chondrogenic differentiation protocols, and angiogenesis models where post-translational fidelity directly influences biological readout. Purity exceeding 95% by SDS-PAGE and endotoxin below 1.0 EU/μg align with standards expected in primary cell culture systems and in vivo regenerative medicine studies where contaminant sensitivity is high.
There are currently no reviews for this product.

