Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
| Code | CSB-PA333076XA01SVG |
| Size | US$299 |
| Image |
|
| Promotion |  |
| Have Questions? | Leave a Message or Start an on-line Chat |
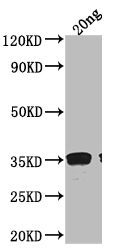
The recombinant Escherichia coli RAD51 protein is administered to a rabbit to trigger an immune response. Antibodies against the RAD51 protein are produced by the rabbit's immune system over time. After an adequate antibody response is generated, a serum sample is collected to obtain polyclonal antibodies. The RAD51 antibody undergoes protein A/G affinity chromatography purification. The effectiveness of the RAD51 antibody is assessed through ELISA and WB applications. This particular RAD51 antibody exhibits specific reactivity with the Escherichia coli RAD51 protein.
The RAD51 protein in Saccharomyces cerevisiae (strain ATCC 204508 / S288c) is a key player in homologous recombination. It mainly facilitates the exchange of genetic material between homologous DNA strands during the repair of double-strand breaks and the restart of stalled or collapsed replication forks. It is crucial for maintaining genome integrity and preventing the accumulation of mutations
There are currently no reviews for this product.
KEGG: sce:YER095W
STRING: 4932.YER095W

