Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
Now, as the Claudins continue to be explored, interest in emerging Claudins is growing fast! Claudins are the key constituent proteins of tight junction (TJ) proteins. TJ proteins play crucial roles of cell-cell adhesion complexes that maintain cell polarity by forming an intramembrane. Encouragingly, a few Claudin family members have been unraveled to make significant impact on tumor pathogenesis, diagnosis, and treatment, like CLDN4, CLDN6, CLDN9, CLDN18.2 etc.
Notably, there have also been a number of studies on Claudin-3. These studies suggest that the abnormal expression of Claudin-3 might take the largely part for the occurrence and development of tumors. Since then, Claudin-3 is expected to be the next new drug target of Claudins family!
1. What's the Tight Junction Protein?
3. What's the Ligand of CLDN3?
Tight junction proteins are composed of integral membrane proteins, membrane-associated, and soluble cytoplasmic proteins. The function of tight junctions is to adhere two cells together fully to prevent the movement of molecules and create a barrier. Importantly, tight junction proteins comprise more than 40 molecules derived from three integral transmembrane protein families: Occludin, Claudins, Junctional Adhesion Molecules (JAMs) [1].
Among them, Claudin-based tight junctions are crucial for the mammalian epidermal barrier, which determine the barrier properties of this type of cell-cell contact existing between the plasma membranes of two neighbouring cells, such as occurring in endothelia or epithelia. It is now widely accepted that loss of intercellular adhesion contributes to the disruption of tight cell junctions, which is linked to the infiltration and metastasis of tumor cells. Given that, Claudin family is considered as a highly sought-after therapeutic target [2].
The tight junction protein 3 (Claudin3/CLDN3) belongs to the family of transmembrane tight junction proteins. CLDN3, with a relative molecular mass of 20 -34 kDa, is the most important backbone protein in tight junctions. Claudins consist of N-terminal and C-terminal, two extracellular loops, and four transmembrane structural domains. It forms a C-terminal PDZ-binding motif that interacts with ZO1, ZO2, ZO3, and multi-PDZ domain protein 1 (MUPP1) (Figure 1) [3].
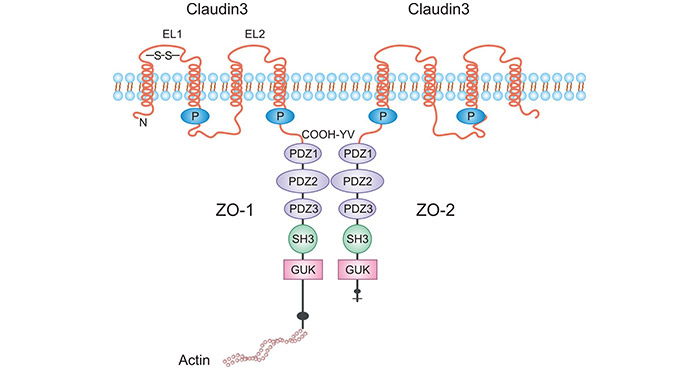
Figure 1. The Structure of CLDN3 [3]
Most studies indicate that CLDN3 is a barrier-forming protein, like blood-brain barrier, the intestinal barrier, and the blood-testis barrier [4-6]. Furthermore, it was also found to modulate cell proliferation, migration, differentiation, and polarity. Of note, CLDN3 expression is dysregulated in a variety of conditions. But it is overexpressed, among others, in ovarian, breast, lung, gastric, and colon cancers [4-6]. Albeit the causal relationship between altered CLDN3 levels and pathogenesis remains to be firmly established, it is likely that CLDN3 impacts key downstream outcomes [4-6].
Clostridium perfringens enterotoxin (CPE) has been identified as a high-affinity natural ligand for CLDN3. Clostridium perfringens enterotoxin (CPE) triggers lysis of epithelial cells through binding to CLDN3 (Figure 2) [7-8]. According to many studies, CLDN4 is overexpressed in many epithelial malignancies and it has been correlated with cancer progression. In addition to CLDN3, clostridium perfringens enterotoxin is also a natural ligand for the CLDN4 protein. Thus, the binding of CPE to CLDN3 and CLDN4 proteins can lead to rapid cell lysis [9].
Recently, CPE is regarded as a therapeutic tool for epithelial malignant tumors by utilizing the damage of CLDN4 and CLDN3. For example, in pancreatic cancer cells, where CLDN-3 and -4 are highly expressed, intra-tumor injection of xenografted pancreatic cancer cells using cytopathic effect (CPE) resulted in extensive necrosis of tumor cells and significantly retarded tumor growth [10]
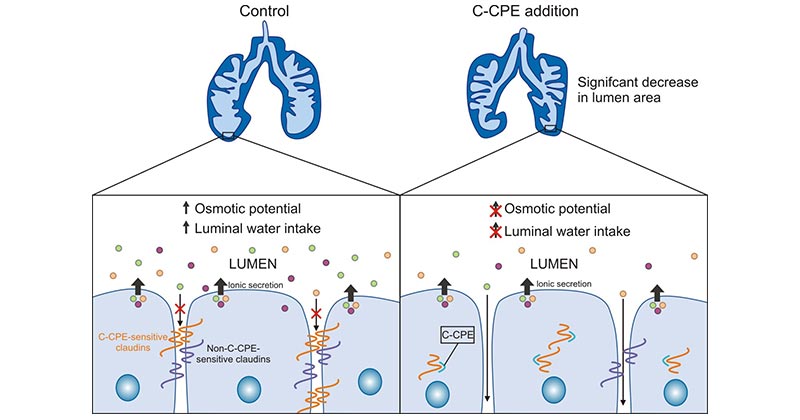
Figure 2. CLDN3 binding to CPE promotes cytolytic necrosis [8]
It was hypothesized that CLDN3 gene expression may contribute to the pathogenesis of tumors in a tissue-specific manner, which further influence the intracellular molecular signaling pathways. However, the exact mechanism of the relationship between aberrant CLDN3 expression and tumors remains incompletely characterized. In hepatocellular carcinoma, CLDN3 is able to downregulate glycogen synthase kinase by GSK3B, CTNNB1, SNAI2 and CDH2, leading to the Wnt/β-catenin-EMT metastatic axis inactivation, thereby significantly inhibiting the metastasis of hepatocellular carcinoma [11-12]. Besides, CLDN3 knockdown also decreased the expression of related bile acid metabolism genes in the liver (like Cyp27a1, Ces1b and Akr1c6) [13]. In addition, loss of CLDN3 or CLDN4 activates the PI3K pathway by activating Akt phosphorylation, resulting in increased expression levels of the PI3K, suggesting that their deletion can promote epithelial mesenchymal transformation (EMT) process [14].
In lung cancer, CLDN3 was significantly overexpressed. Further studies suggested CLDN3 overexpression was associated with ERK1/2 and PI3K-Akt signaling pathways [14]. In colorectal cancer tissues, CLDN3 in CRC tissues can be elevated by activation of the SCF/c-kit/JNK/AP-1 signaling pathway [15]. In ovarian cancer, CLDN3 expression was affected by ZDHHC12, implying that targeting ZDHHC12-mediated CLDN3 S-palmitoylation may be a potential strategy for ovarian cancer treatment (Figure 3) [16]. Additional studies have indicated that CLDN3 may inhibit growth and metastatic of cells via maintaining E-cadherin expression and limiting the expression of β-cadherin [17].
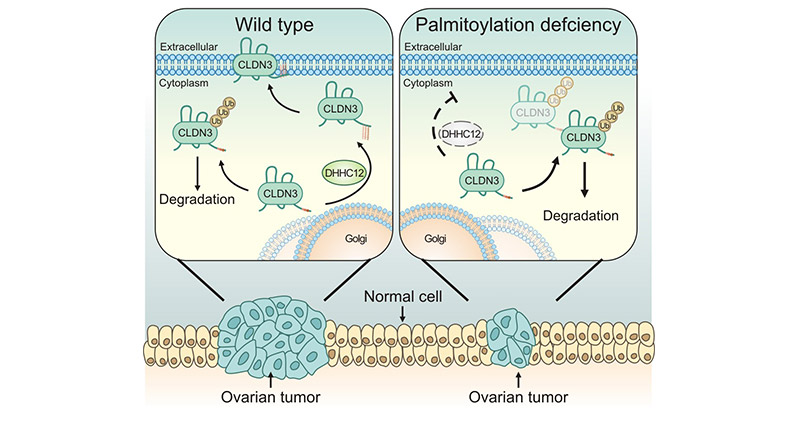
Figure 3. CLDN3 is regulated by palmitoyltransferase ZDHHC12 [16]
CLDN3, as a key sealing claudin, constitutes the tight junctions of endothelial cells. But CLDN3 abnormal expression can lead to damage to the intercellular structure of endothelial cells. Disruption of tight junctions may lead to weakened cell adhesion, resulting in loss of tumor cell polarity, uncontrolled growth, and even tumor invasion and metastasis. CLDN3 has emerged as modulators of proliferation, migration, and differentiation. Such effects might be the key during development and in a variety of diseases.
In colorectal cancer, epidermal growth factor (EGF) increased the expression of CLDN3 in colorectal cancer cell line HT-29 cells and promoted cell migration, suggesting that overexpressed CLDN3 enhances the invasive potential of colorectal cancer cells [14]. Typically, EGF is a marker in the development of certain tumors. EGF receptor antagonists such as Gefitinib have been used in clinical anti-tumor therapy.
In prostate cancer, CLDN3 was upregulated in prostate hyperplastic tissue. Using northern blot analysis, significantly higher levels of CLDN3 were found in human prostate cancer tissues compared to surrounding normal tissues [18]; the same was true in nasopharyngeal carcinoma, where CLDN3 expression was higher in nasopharyngeal carcinoma tissue than in nasopharyngeal paraneoplastic tissue. Plus, CLDN3 expression was higher in nasopharyngeal carcinoma tissues with higher pathological stage [19].
In cervical cancer Hela cell lines, SST promotes the CLDN3 and CLDN4 expression, thereby inhibiting the development of cervical cancer [20-22]. In breast cancer, CLDN3 was positively correlated with BRCA1-mutated BC [23]. In squamous epithelial cell carcinoma, decreased CLDN3 expression was significantly associated with tumor TNM stage, lymph node metastasis, and tumor recurrence [24-25]. In hepatocellular carcinoma, CLDN3 was significantly associated with survival of patients with hepatocellular carcinoma. These studies suggested that CLDN3 could be further evaluated as a novel biomarker or therapeutic target for different tumors [5-6].
Blood-brain barrier (BBB) disruption is thought to play a critical role in the pathophysiology of ischemia/reperfusion. CLDN3 is closely related to the function of the BBB and can alter the permeability of the BBB [26]; Gallstone disease is most common liver disorders in adults. In the liver of CLDN3-knockout mice, the paracellular phosphate ion permeability through hepatic tight junctions was significantly increased, resulting in calcium phosphate core formation. Cholesterol overdose could cause cholesterol gallstone disease in these mice [27]. In fact, CLDN3, as the most expressed TJ protein in the liver, such as intestinal dysbiosis, chronic liver disease, even malignant liver tumors [28].
CLDN3 is an important component of the CLDN family and serves a substantial role in maintaining the function of TJs, which is involved in the selective permeation and cell polarization of tight junctions. In particular, due to its high tissue specificity, CLDN3 has been widely studied and plays an important role in the development and progression of many tumors. Data from Pharmsnap show that several preclinical and drug discovery stage CLDN3-related drugs are already in development (Table 1). Currently, CLDN3-based targeted therapies are emerging as an important research direction for the CLDN family, holding significant drug discovery potential!
| Drugs | Target | Mechanism | Drug Type | Indications | Institutes | R&D status |
|---|---|---|---|---|---|---|
| ABN 501;ABN-501;Anti-CLDN3 monoclonal antibody - Abion | CLDN3 | CLDN3 inhibitor | Monoclonal antibodies | Ovarian Cancer | ABION, Inc. | Preclinical |
| KM-3907;dual claudin-3/claudin-4 mAb (Kyowa Hakko Kirin) | CLDN3;CLDN1 |
CLDN3 inhibitor; CLDN1 inhibitor |
Bispecific antibodies | / | / | Drug Discovery |
| MORAb-075;anti-claudin-3/4 humanized mAb(Morphotek) | CLDN3; CLDN1 |
CLDN3 inhibitor; CLDN1 inhibitor |
Monoclonal antibodies | / | / | Drug Discovery |
Table 1: Clinical drugs for CLDN3
To fully support researchers and pharmaceutical companies in their research on CLDN3 in tumor and other diseases, CUSABIO presents the CLDN3 active protein (Code: CSB-MP005505HU) to support your research on the mechanism of CLDN3 or its potential clinical value.
Recombinant Human Claudin-3(CLDN3)-VLPs (Active)

The high specifity was validated by western blot. (Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
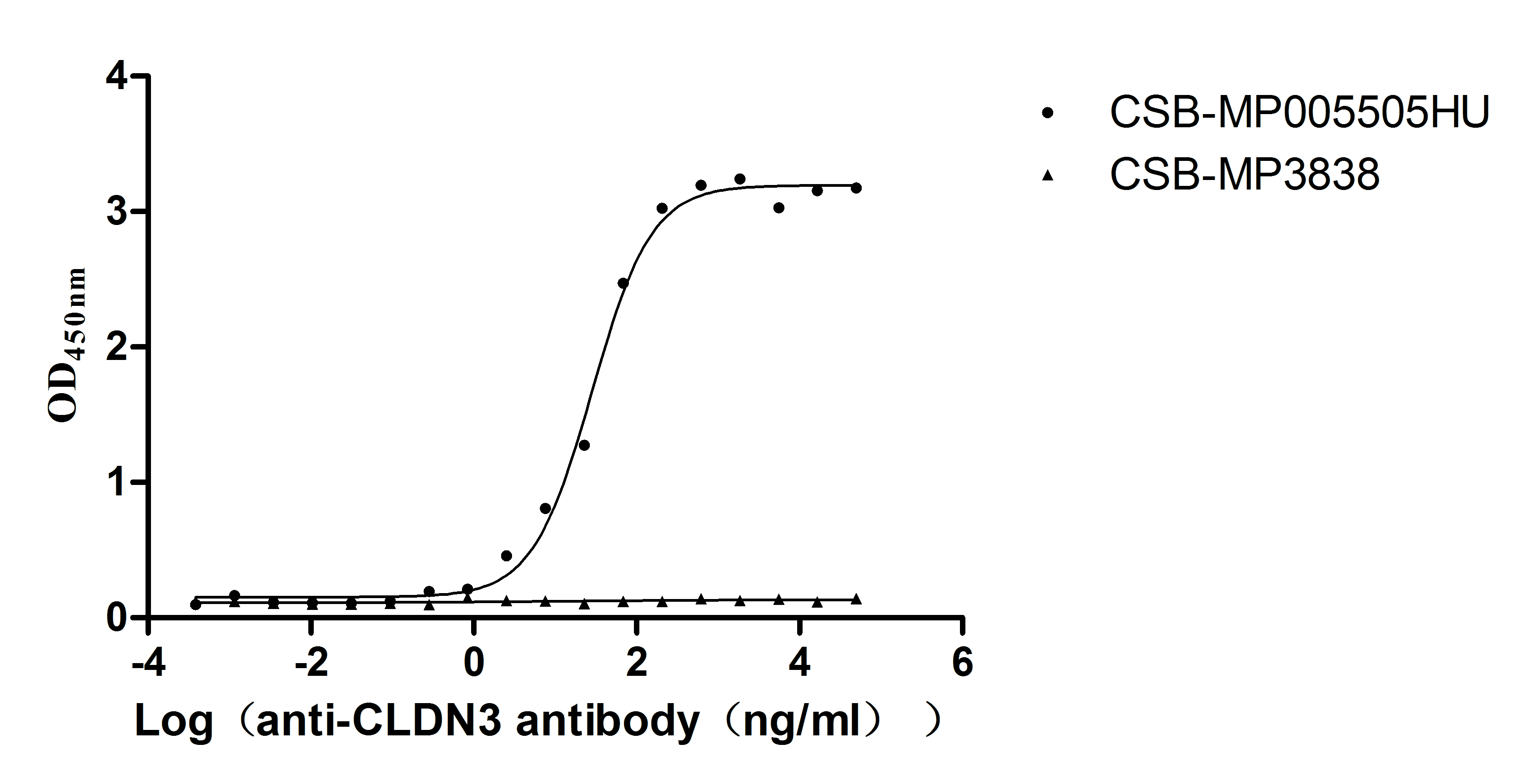
Immobilized Human CLDN3 at 10μg/mL can bind Anti-CLDN3 recombinant antibody (CSB-RA005505MA1HU), the EC50 is 23.62-34.37 ng/mL.VLPs (CSB-MP3838) is negative control.
References
[1] Fujiwara, Sachiko, et al. "Tight junction formation by a claudin mutant lacking the COOH-terminal PDZ domain-binding motif ." Annals of the New York Academy of Sciences 1516.1 (2022): 85-94.
[2] Castro Dias, Mariana, et al. "Claudin-3-deficient C57BL/6J mice display intact brain barriers." scientific reports 9.1 (2019): 203.
[3] Che, J., Yue, D., et al. "Claudin-3 Inhibits Lung Squamous Cell Carcinoma Cell Epithelial-mesenchymal Transition and Invasion via Suppression of the Wnt/β-Catenin Signaling Pathway." International Journal of Medical Sciences, 15(4) (2018): 339-351.
[4] Lei, Ningjing, et al. "Claudin-3 inhibits tumor-induced lymphangiogenesis via regulating the PI3K signaling pathway in lymphatic endothelial cells. " Scientific Reports 12.1 (2022): 17440.
[5] Wang, ZhaoHan, et al. "Plasma claudin-3 is associated with tumor necrosis factor-alpha-induced intestinal endotoxemia in liver disease." Clinics and Research in Hepatology and Gastroenterology 43.4 (2019): 410-416.
[6] Yang, Y., et al. "Mo1775 Epigenetic Inactivation of Claudin 3 in Hepatocellular Carcinoma and Its Functional Consequences." Gastroenterology 5.144 ( 2013): S-1023.
[7] Torres, Julia Baguña, et al. "Radiolabeled cCPE peptides for SPECT imaging of claudin-4 overexpression in pancreatic cancer." journal of Nuclear Medicine 61.12 (2020): 1756-1763.
[8] La Charité-Harbec, Simon, et al. "Claudin-3 regulates luminal fluid accumulation in the developing chick lung." Differentiation 124 (2022): 52-59.
[9] Li, Xiangru, et al. "Development of an Anti-Claudin-3 and -4 Bispecific Monoclonal Antibody for Cancer Diagnosis and Therapy." journal of Pharmacology and Experimental Therapeutics 351.1 (2014): 206-213.
[10] Pahle, Jessica, et al. "Effective oncoleaking treatment of pancreatic cancer by claudin-targeted suicide gene therapy with Clostridium perfringens enterotoxin (CPE)." Cancers 13.17 (2021): 4393.
[11] Gasperoni, Jemma G., et al. "Grainyhead-like (Grhl) target genes in development and cancer." International Journal of Molecular Sciences 23.5 (2022): 2735.
[12] Ahmad, Rizwan, et al. "Loss of claudin-3 expression induces IL6/gp130/Stat3 signaling to promote colon cancer malignancy by hyperactivating Wnt/β- catenin signaling." Oncogene 36.47 (2017): 6592-6604.
[13] Reiling, Janske, et al. "Low-Dose Lipopolysaccharide Causes Biliary Injury by Blood Biliary Barrier Impairment in a Rat Hepatic Ischemia /Reperfusion Model." Liver Transplantation 23.2 (2017): 194-206.
[14] de Souza, Waldemir F., et al. "Claudin-3 overexpression increases the malignant potential of colorectal cancer cells: roles of ERK1/2 and PI3K-Akt as modulators of EGFR signaling." ploS one 8.9 (2013): e74994.
[15] Wang, Yaxi, et al. "SCF/C-Kit/JNK/AP-1 signaling pathway promotes claudin-3 expression in colonic epithelium and colorectal carcinoma." International journal of molecular sciences 18.4 (2017): 765.
[16] Yuan, Meng, et al. "ZDHHC12-mediated claudin-3 S-palmitoylation determines ovarian cancer progression." Acta Pharmaceutica Sinica B 10.8 (2020): 1426-1439.
[17] Che, Juanjuan, et al. "Decreased expression of claudin-3 is associated with a poor prognosis and EMT in completely resected squamous cell lung carcinoma ." Tumor Biology 36 (2015): 6559-6568.
[18] Orea, María J., et al. "Claudin-3 Loss of Expression Is a Prognostic Marker in Castration-Resistant Prostate Cancer." International Journal of Molecular Sciences 24.1 (2023): 803.
[19] Tessier-Cloutier, Basile, et al. "Proteomic analysis of transitional cell carcinoma-like variant of tubo-ovarian high-grade serous carcinoma." Human pathology 101 (2020): 40-52.
[20] Chen, Peng, et al. "Structural basis for CSPG4 as a receptor for TcdB and a therapeutic target in Clostridioides difficile infection." Nature communications 12.1 (2021): 3748.
[21] Xie, Xiaoxi, et al. "Roles of gastrointestinal polypeptides in intestinal barrier regulation." Peptides (2022): 170753.
[22] Liu, Jinhui, et al. "Development of an immune gene prognostic classifier for survival prediction and response to immunocheckpoint inhibitor therapy/ chemotherapy in endometrial cancer." International immunopharmacology 86 (2020): 106735.
[23] Szade, Jolanta, et al. "Comparison of claudin-3 and claudin-4 expression in bilateral and unilateral breast cancer." Neoplasma 68.2 (2020): 283-289.
[24] Feng, Ji, et al. "Capsaicin inhibits migration and invasion via inhibiting epithelial-mesenchymal transition in esophageal squamous cell carcinoma by up-regulation of claudin-3 expression." Journal of Functional Foods 89 (2022): 104934.
[25] Che, Juanjuan, et al. "Claudin-3 inhibits lung squamous cell carcinoma cell epithelial-mesenchymal transition and invasion via suppression of the Wnt /β-catenin signaling pathway." international journal of medical sciences 15.4 (2018): 339.
[26] Haseloff, Reiner F., et al. "Transmembrane proteins of the tight junctions at the blood-brain barrier: structural and functional aspects ." Seminars in cell & developmental biology. vol. 38. Academic Press, 2015.
[27] Tanaka, Hiroo, et al. "Claudin-3 regulates bile canalicular paracellular barrier and cholesterol gallstone core formation in mice." journal of hepatology 69.6 (2018): 1308-1316.
[28] Baier, Felix Alexander, et al. "Loss of Claudin-3 Impairs Hepatic Metabolism, Biliary Barrier Function, and Cell Proliferation in the Murine Liver." Cellular and molecular gastroenterology and hepatology 12.2 (2021): 745-767.


Comments
Leave a Comment