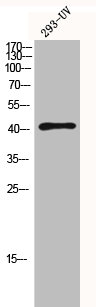
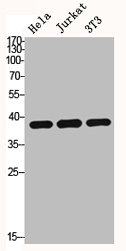
Alternative Names
CDA07 antibody; CDLS5 antibody; HD 8 antibody; HD8 antibody; HDAC 8 antibody; Hdac8 antibody; HDAC8_HUMAN antibody; HDACL 1 antibody; HDACL1 antibody; Histone deacetylase 8 antibody; Histone deacetylase like 1 antibody; MRXS6 antibody; RPD 3 antibody; RPD3 antibody; WTS antibody
Species Reactivity
Human,Mouse,Rat,Monkey
Immunogen
Synthesized peptide derived from Human HDAC8 around the non-phosphorylation site of S39.
Immunogen Species
Homo sapiens (Human)
Purification Method
The antibody was affinity-purified from rabbit antiserum by affinity-chromatography using epitope-specific immunogen.
Concentration
It differs from different batches. Please contact us to confirm it.
Buffer
Liquid in PBS containing 50% glycerol, 0.5% BSA and 0.02% sodium azide.
Tested Applications
WB, IHC, ELISA
Recommended Dilution
| Application |
Recommended Dilution |
| WB |
1:500-1:2000 |
| IHC |
1:100-1:300 |
| ELISA |
1:20000 |
Storage
Upon receipt, store at -20°C or -80°C. Avoid repeated freeze.
Lead Time
Basically, we can dispatch the products out in 1-3 working days after receiving your orders. Delivery time maybe differs from different purchasing way or location, please kindly consult your local distributors for specific delivery time.
Usage
For Research Use Only. Not for use in diagnostic or therapeutic procedures.