
Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
SLC3A2 (CD98hc) is far more than a typical transporter protein; it serves as a pivotal hub linking amino acid metabolism to cellular signaling. It plays a critical role in cancer, neurological, metabolic, and immune diseases, and is emerging as a highly promising therapeutic target. Following this thread, this article will elucidate its comprehensive pathway from amino acid transport mechanisms to its development as an emerging therapeutic target.
1. Why Is SLC3A2 a Central Hub in Metabolism and Disease?
2. How Does SLC3A2 Execute Its Key Biological Functions?
3. How Does SLC3A2 Regulate Critical Cellular Signaling Pathways?
4. In Which Major Diseases Does SLC3A2 Play a Key Role?
5. What Is the Progress in Developing Drugs Targeting SLC3A2?
SLC3A2 (Solute Carrier Family 3 Member 2, also known as 4F2hc or CD98hc) is a type II transmembrane glycoprotein belonging to the SLC3 family. Its primary function is to form heterodimeric amino acid transporters (HATs) with light chain subunits from the SLC7 family, jointly mediating the transmembrane transport of amino acids and regulating cellular metabolism and signaling activities [1,2].
SLC3A2 consists of a short N-terminal cytoplasmic region, a single transmembrane helix (TM), and a large C-terminal extracellular domain (ED). Although structurally similar to bacterial glucosidases, the extracellular domain lacks catalytic activity and primarily functions in protein-protein interactions and conformational stabilization [1]. Via conserved disulfide bonds, SLC3A2 forms complexes with SLC7 family members (such as LAT1, xCT, etc.) to fulfill its transport function. In this heterodimer, SLC3A2 is responsible for trafficking the light chain subunits to the plasma membrane and maintaining their stability, while substrate recognition and ion selectivity are determined by the light chain subunits [2,3].
SLC3A2 is highly expressed in various tissues with high metabolic rates, including the blood-brain barrier (BBB), placenta, and immunologically activated cells [4-7]. Its absence during embryonic development is lethal, while in tumor cells, SLC3A2 is often significantly upregulated to enhance amino acid supply and promote growth and metabolic reprogramming [8,9]. These characteristics establish SLC3A2 as a crucial hub connecting nutrient metabolism and signal regulation.
SLC3A2 mediates the transport of various types of amino acids by forming complexes with different light chain subunits:
N-glycosylation is a key modification for maintaining SLC3A2 function. In pancreatic cancer, glycosylation at Asn365, catalyzed by B3GNT3, significantly enhances protein stability and xCT binding, thereby sustaining system xc⁻ activity and protecting against ferroptosis [9]. Defects in glycosylation accelerate degradation and increase sensitivity to ferroptosis.
SLC3A2 is primarily localized to the plasma membrane but can also be targeted to the lysosomal membrane due to signaling stimuli or protein interactions (e.g., with LAPTM4b or DRAM-1), participating in amino acid storage and mTORC1 signaling regulation [10,15].
SLC3A2-mediated leucine uptake is a key step for activating the mTORC1 signaling pathway. Leucine regulates Rag GTPase activity through the Sestrin-GATOR and LRS pathways, promoting mTORC1 activation on the lysosomal membrane, thereby stimulating protein synthesis and cell growth [16,17]. In head and neck squamous cell carcinoma, loss of SLC3A2 reduces mTORC1 activity and triggers autophagy, while inhibition of autophagy enhances radiosensitivity [3].
When SLC3A2 function is impaired, amino acid depletion leads to the accumulation of uncharged tRNA, activating the GCN2–eIF2α–ATF4 pathway. ATF4, in turn, promotes the expression of SLC3A2 and LAT1, restoring amino acid homeostasis [19]. This mechanism is particularly prominent in high-metabolism tumors such as prostate cancer.
The intracellular domain of SLC3A2 can interact with integrin β subunits, influencing cell adhesion and migration [2,20]. In liver cancer, SLC3A2 can inhibit β1 integrin activation to restrict invasion; whereas in renal cancer, it promotes integrin signaling, enhancing migration and matrix adhesion capabilities [20].
As a nodal point linking amino acid metabolism and signaling pathways, SLC3A2 is closely associated with the occurrence and progression of various diseases.
SLC3A2 is a downstream target gene of the androgen receptor splice variant AR-V7. AR-V7 drives its expression and activates mTORC1 signaling via the LAT1 complex, thereby supporting the growth of castration-resistant prostate cancer (CRPC) [8,11]. Clinical data show that high SLC3A2 expression is closely correlated with higher pathological grade and poor prognosis [21].
In pancreatic ductal adenocarcinoma (PDAC), B3GNT3-mediated N-glycosylation enhances SLC3A2 stability and promotes xCT binding, maintaining system xc⁻ function and antioxidant capacity [9]. This mechanism confers significant tolerance to ferroptosis in PDAC cells. Analysis of clinical samples indicates that dual high expression of SLC3A2 and B3GNT3 is closely associated with shortened survival, suggesting their potential as prognostic biomarkers.
High SLC3A2 expression promotes amino acid transport and DNA repair capacity, conferring radioresistance to tumor cells [3]. Knockdown of SLC3A2 or inhibition of mTORC1 can attenuate radioresistance, and combined autophagy inhibition further enhances therapeutic efficacy, suggesting its potential as a target for radiosensitization.
In bladder cancer and renal cell carcinoma, the SLC3A2–LAT1 complex mediates leucine uptake, promoting cell proliferation and invasion [21,22]. High SLC3A2 expression is closely associated with high grade, metastatic risk, and poor prognosis, indicating its potential diagnostic and therapeutic value in urological tumors.
In hepatocellular carcinoma, breast cancer, and glioma, SLC3A2 has also been found to regulate the mTORC1 pathway and integrin signaling, thereby influencing cell metabolism, migration, and stress response capacity. These findings support its potential as a dual "metabolism-signaling" target in precision cancer therapy.
SLC3A2 is highly expressed at the blood-brain barrier (BBB) and is closely involved in maintaining brain amino acid homeostasis. LAT1 gene mutations (Ala246Val, Pro375Leu) impair the function of the SLC3A2-LAT1 complex, leading to brain amino acid imbalance, activation of the ISR pathway, and phenotypes associated with autism spectrum disorder (ASD) [22,23]. Furthermore, in Parkinson's disease, the SLC3A2–LAT1 complex is responsible for the brain transport of L-DOPA. Branched-chain amino acids (BCAAs) can competitively bind to it, reducing L-DOPA treatment efficiency, explaining the drug tolerance observed in some patients [24].
SLC3A2 is closely linked to BCAAs metabolism, insulin resistance, and type 2 diabetes (T2DM). BCAA uptake mediated by the SLC3A2-LAT1 complex can excessively activate mTORC1, thereby inhibiting insulin signaling and inducing insulin resistance [25]. Additionally, hyperglycemia can inhibit AMPK signaling and downregulate SLC3A2 expression, further elevating plasma BCAA levels and creating a vicious metabolic cycle [25]. In pancreatic β-cells, SLC3A2 participates in amino acid uptake and insulin synthesis; its dysfunction can lead to insufficient secretion and exacerbate metabolic imbalance [26].
SLC3A2 plays a key role in the metabolic reprogramming of immune cells.
These results indicate that SLC3A2 is not only a metabolic channel protein but also participates in the metabolic regulation of immune signals, representing a new potential therapeutic target for inflammatory diseases.
SLC3A2 has emerged as a novel anti-cancer drug target. Current research directions cover monoclonal antibodies, small molecule inhibitors, and antibody-drug conjugates (ADCs), primarily focused on hematological malignancies, solid tumors, and brain cancers, all currently in the preclinical stage.
| Drug | Mechanism of Action | Drug Type | Indications Under Investigation | Highest R&D Stage |
|---|---|---|---|---|
| MAb52-4.2 | SLC3A2 Modulator | Monoclonal Antibody | Hematological Tumors | Solid Tumors | Preclinical |
| 211At-TLX-102 | SLC3A2 Inhibitor | SLC7A5 Inhibitor | Small Molecule / Therapeutic Radiopharmaceutical | Brain Cancer | Preclinical |
| c-SF-25 Mab(Kagoshima University) | SLC3A2 Inhibitor | Monoclonal Antibody | Adult T-cell Leukemia/Lymphoma | Preclinical |
| NPB15 | SLC3A2 Inhibitor | Monoclonal Antibody | Hepatocellular Carcinoma | Preclinical |
| HH018-sesutecan | SLC3A2 Inhibitor | ADC | Tumors | Preclinical |
(Data current as of October 30, 2025, sourced from Synapse)
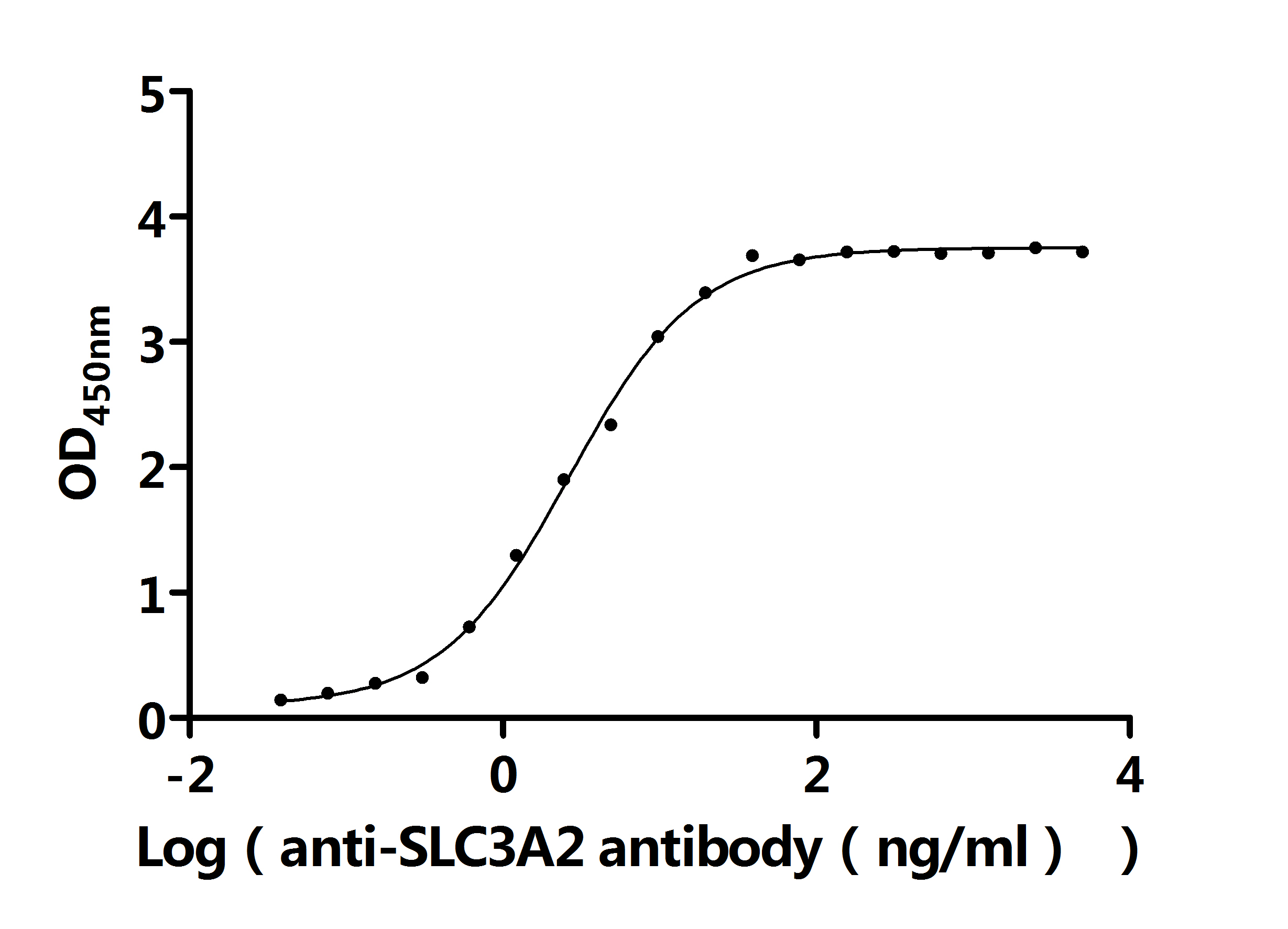
● SLC3A2 Recombinant Protein
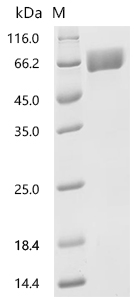
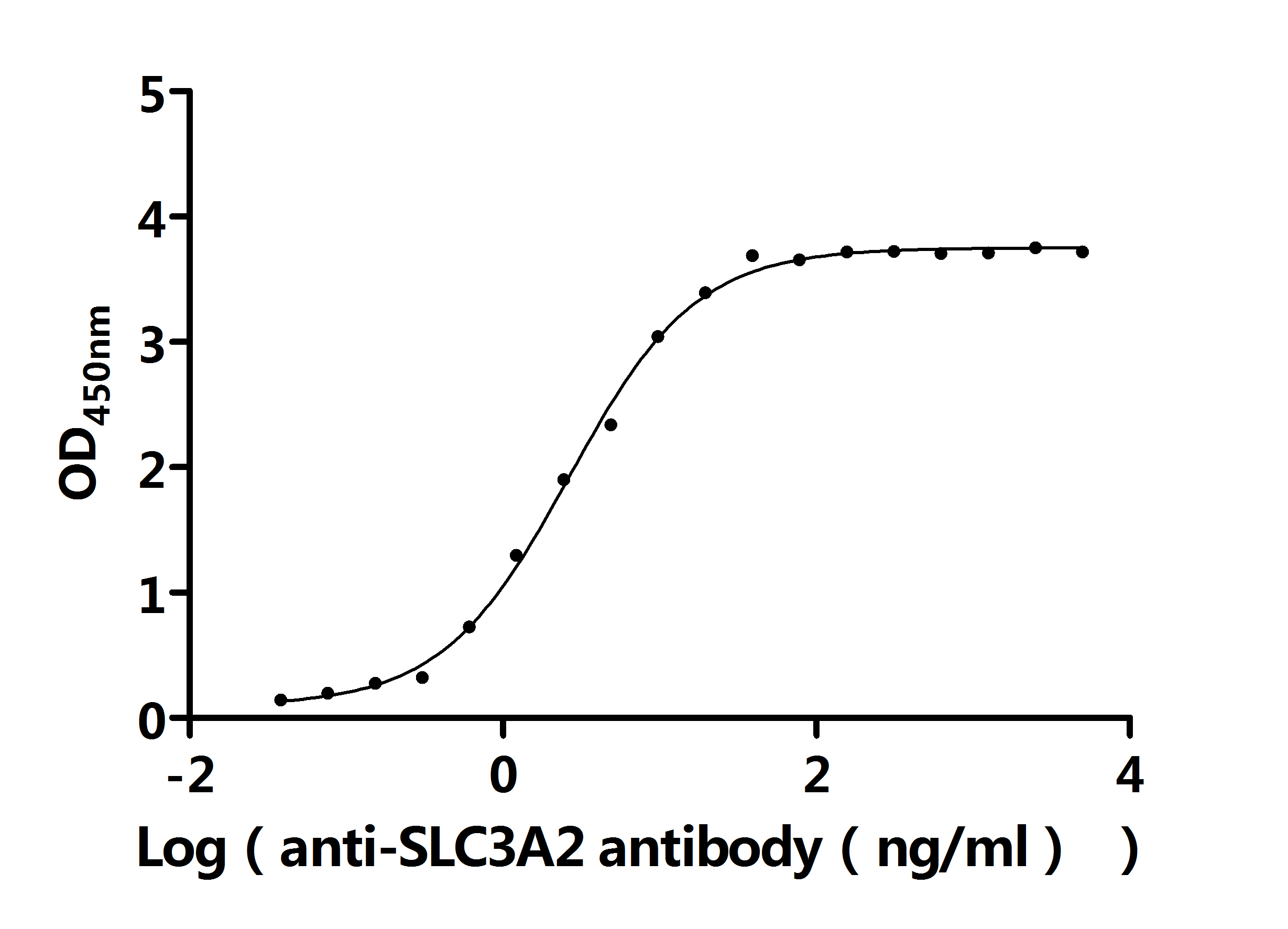
● SLC3A2 Antibodies
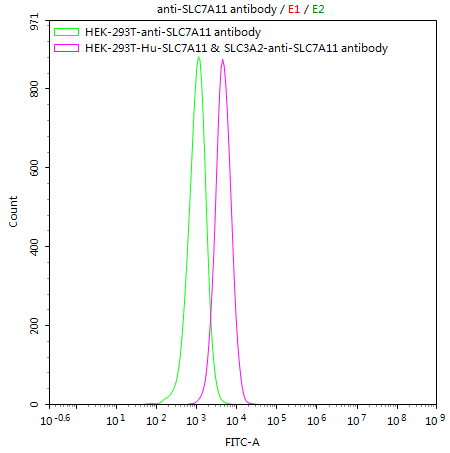
● SLC3A2 Cell Line
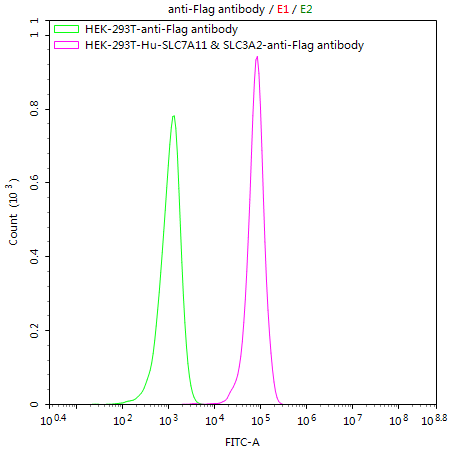
HEK293T/Human SLC7A11 & SLC3A2 Stable Cell Line; CSB-SC5338HU
References
[1] Sub-Nanometer Cryo-EM Density Map of the Human Heterodimeric Amino Acid Transporter 4F2hc-LAT2.
[2] Kahlhofer J, Teis D. The human LAT1–4F2hc (SLC7A5–SLC3A2) transporter complex: Physiological and pathophysiological implications[J]. Basic Clin Pharmacol Toxicol, 2023, 133(5):459-472.
[3] Digomann D, Linge A, Dubrovska A. SLC3A2/CD98hc, autophagy and tumor radioresistance: a link confirmed[J]. Autophagy, 2019, 15(10):1850-1851.
[4] Boado RJ, Li JY, Nagaya M, et al. Selective expression of the large neutral amino acid transporter at the blood–brain barrier[J]. Proc Natl Acad Sci USA, 1999, 96(21):12079-12084.
[5] Matsuo H, Tsukada S, Nakata T, et al. Expression of a system L neutral amino acid transporter at the blood–brain barrier[J]. Neuroreport, 2000, 11(16):3507-3511.
[6] Laudicella R, Albano D, Alongi P, et al. (18)F-Facbc in Prostate Cancer: A Systematic Review and Meta-Analysis[J]. Cancers, 2019, 11(1348).
[7] Nii T, Segawa H, Taketani Y, et al. Molecular events involved in up-regulating human Na+-independent neutral amino acid transporter LAT1 during T-cell activation[J]. Biochem J, 2001, 358(Pt 3):693-704.
[8] Zhao X, Sakamoto S, Maimaiti M, et al. Contribution of LAT1-4F2hc in Urological Cancers via Toll-like Receptor and Other Vital Pathways[J]. Cancers, 2022, 14(1):229.
[9] Ma H, Chen X, Mo S, et al. Targeting N-glycosylation of 4F2hc mediated by glycosyltransferase B3GNT3 sensitizes ferroptosis of pancreatic ductal adenocarcinoma[J]. Cell Death Differ, 2023, 30(1988-2004).
[10] Helgudóttir S-S, Johnsen K-B, Routhe L-G, et al. Upregulation of Transferrin Receptor 1 (TfR1) but Not Glucose Transporter 1 (GLUT1) or CD98hc at the Blood–Brain Barrier in Response to Valproic Acid[J]. Cells, 2024, 13(14):1181.
[11] Sugiura M, Sato H, Okabe A, et al. Identification of AR-V7 downstream genes commonly targeted by AR/AR-V7 and specifically targeted by AR-V7 in castration resistant prostate cancer[J]. Transl Oncol, 2021, 14(10):100915.
[12] Torrents D, Estévez R, Pineda M, et al. Identification and characterization of a membrane protein (y+L amino acid transporter-1) that associates with 4F2hc to encode the amino acid transport activity y+L[J]. J Biol Chem, 1998, 273(49):32437-32445.
[13] Lee Y, Wiriyasermkul P, Jin C, et al. Cryo-EM structure of the human L-type amino acid transporter 1 in complex with glycoprotein CD98hc[J]. Nat Struct Mol Biol, 2019, 26(6):510-517.
[14] Deuschle F-C, Schiefner A, Brandt C, et al. Design of a surrogate Anticalin protein directed against CD98hc for preclinical studies in mice[J]. Protein Sci, 2020, 29(1774-1783).
[15] Brown EJ, Albers MW, Shin TB, et al. A mammalian protein targeted by G1-arresting rapamycin-receptor complex[J]. Nature, 1994, 369(6483):756-758.
[16] Saxton RA, Knockenhauer KE, Wolfson RL, et al. Structural basis for leucine sensing by the Sestrin2-mTORC1 pathway[J]. Science, 2016, 351(6268):53-58.
[17] Han JM, Jeong SJ, Park MC, et al. Leucyl-tRNA synthetase is an intracellular leucine sensor for the mTORC1-signaling pathway[J]. Cell, 2012, 149(2):410-424.
[18] Park Y, Reyna-Neyra A, Philippe L, et al. mTORC1 balances cellular amino acid supply with demand for protein synthesis through post-transcriptional control of ATF4[J]. Cell Rep, 2017, 19(6):1083-1090.
[19] Cordova RA, Misra J, Amin PH, et al. GCN2 eIF2 kinase promotes prostate cancer by maintaining amino acid homeostasis[J]. Elife, 2022, 11:e81083.
[20] Poettler M, Unseld M, Braemswig K, et al. CD98hc (SLC3A2) drives integrin-dependent renal cancer cell behavior[J]. Mol Cancer, 2013, 12(1):169.
[21] Betsunoh H, Fukuda T, Anzai N, et al. Increased expression of system large amino acid transporter (LAT)-1 mRNA is associated with invasive potential and unfavorable prognosis of human clear cell renal cell carcinoma[J]. BMC Cancer, 2013, 13(509).
[22]Tarlungeanu DC, Deliu E, Dotter CP, et al. Impaired amino acid transport at the blood brain barrier is a cause of autism spectrum disorder[J]. Cell, 2016, 167(6):1481-1494.
[23] Errasti-Murugarren E, Palacín M. Heteromeric amino acid transporters in brain: from physiology to pathology[J]. Neurochem Res, 2022, 47(1):23-36.
[24] Beckers M, Bloem BR, Verbeek MM. Mechanisms of peripheral levodopa resistance in Parkinson’s disease[J]. NPJ Parkinsons Dis, 2022, 8(1):56.
[25] Yamamoto Y, Sawa R, Wake I, et al. Glucose-mediated inactivation of AMP-activated protein kinase reduces the levels of L-type amino acid transporter 1 mRNA in C2C12 cells[J]. Nutr Res, 2017, 47(13-20).
[26] Kobayashi N, Okazaki S, Sampetrean O, et al. CD44 variant inhibits insulin secretion in pancreatic β cells by attenuating LAT1-mediated amino acid uptake[J]. Sci Rep, 2018, 8(1):2785.
[27] Hayashi K, Kaminuma O, Nishimura T, et al. LAT1-specific inhibitor is effective against T cell-mediated allergic skin inflammation[J]. Allergy, 2020, 75(2):463-467.
[28] Torigoe M, Maeshima K, Ozaki T, et al. L-leucine influx through Slc7a5 regulates inflammatory responses of human B cells via mammalian target of rapamycin complex 1 signaling[J]. Mod Rheumatol, 2019, 29(5):885-891.


Comments
Leave a Comment