Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
Tumor drug resistance and metastasis are major causes of cancer treatment failure and patient mortality. Confronting this clinical challenge, scientists have turned their attention to a critical "signaling hub" on cancer cells – the EphA2 receptor.
Substantial research indicates that EphA2 is not only highly expressed in various cancers but also plays a central role in driving drug resistance and promoting invasion and metastasis. This article will delve into the unique dual signaling mechanisms of EphA2, explain how it acts as an "accomplice" in tumor malignancy, and review the novel targeted therapeutic strategies being developed around it, revealing this promising new focus in the fight against cancer.
1. Research Background on EphA2 Receptor Tyrosine Kinase
2. Molecular Background, Structure, and Basic Functions of EphA
3. EphA2 Signaling Pathways and Research Mechanisms
4. The Role of EphA2 in Related Diseases
5. Advances in EphA2-Targeted Drugs and Therapeutic Strategies
EphA2 (Erythropoietin-producing human hepatocellular receptor A2) is an important member of the Eph receptor tyrosine kinase (RTK) family, the largest known RTK subfamily, playing a central role in cell communication, development, morphogenesis, migration, and adhesion [1,2]. EphA2 activation depends on ligand binding and can form oligomers of different conformations, triggering diverse signaling responses [3], thus having broad implications in both physiological and pathological states.
Extensive research confirms that EphA2 is highly expressed in numerous malignant tumors and is closely associated with tumor aggressiveness, metastasis, and poor prognosis [4-6]. In triple-negative breast cancer, high EphA2 expression is significantly correlated with poorer survival rates, and its gene silencing can inhibit cell proliferation and invasion [4]. Similar pro-cancer effects have been observed in colorectal cancer [5], gastric cancer [7], ovarian cancer [8], non-small cell lung cancer [9], and others. EphA2 can promote tumor cell proliferation, epithelial-mesenchymal transition (EMT) [10], angiogenesis [7,11], metastasis, and drug resistance formation [12,13]. Furthermore, exosomal EphA2 can transfer drug resistance between cells, further exacerbating the malignant process [12].
EphA2 signaling exhibits a typical "dual function," namely ligand-dependent (canonical) and ligand-independent (non-canonical) modes. Canonical signaling often exhibits tumor-suppressive effects; for example, ephrinA1-EphA2 binding can regulate the actin cytoskeleton via Ezrin phosphorylation, promoting cell polarization and adhesion [1,2]. In contrast, in the tumor microenvironment lacking ligand, EphA2 often undergoes phosphorylation at Ser897, forming non-canonical signaling that exhibits pro-oncogenic characteristics [14-16]. For instance, in prostate cancer, loss of androgen receptor (AR) suppression leads to EphA2 overexpression, driving castration resistance via non-canonical signaling [6,15]. Beyond tumors, EphA2 is also involved in inflammation and immune processes, such as LPS-induced lung injury [17] and antiviral responses in nasal mucosa [18,19].
Therefore, due to its critical role in various diseases and signaling complexity, EphA2 has emerged as a challenging yet potential therapeutic target [20,21]. Current research and development actively focus on small-molecule inhibitors, monoclonal antibodies, ADC drugs, and nanodelivery systems targeting EphA2 [22,23]. For example, the inhibitor ALW-II-41-27 can inhibit cervical cancer growth and reverse colorectal cancer drug resistance [5,24]. Additionally, EphA2 combination therapy with PARP or HDAC inhibitors shows synergistic effects in ovarian cancer [25,26]. Thus, a deep understanding of the EphA2 signaling pathway is crucial for the development of targeted therapies and research into resistance mechanisms.
The Eph receptor tyrosine kinase (Eph RTK) family is an important molecular network mediating cell-cell communication, divided into EphA and EphB classes based on sequence homology and ligand specificity. EphA receptors primarily bind GPI-anchored ephrinA ligands, while EphB receptors bind transmembrane ephrinB ligands [8]. Unlike other RTKs, Eph/ephrin signaling relies on direct cell-cell contact, and its activation requires the formation of receptor-ligand complexes across membranes [27]. This structural feature enables EphA2 to play a key role in cell recognition, migration, and morphological changes. For example, EphA2 binding to ephrinA1 can regulate cell adhesion and repulsion [1,2], significant in both development and tumor growth. Different ephrin ligands (such as EFNA1 and EFNA5) may have opposing effects in disease contexts [8], reflecting the highly context-dependent nature of Eph/ephrin signaling.
The EphA2 structure includes an extracellular ligand-binding domain, a transmembrane region, and an intracellular tyrosine kinase domain. The extracellular domain is responsible for recognizing EphrinA ligands, while the intracellular domain contains the juxtamembrane region, kinase domain, and a PDZ-binding motif for recruiting downstream signaling complexes [3]. Its activation is often accompanied by receptor dimerization and autophosphorylation. Studies show that decreased membrane cholesterol can promote EphA2 self-assembly and Ser897 phosphorylation, enhancing its pro-oncogenic signaling [28].
EphA2 is widely expressed in normal tissues, such as vascular smooth muscle cells and sinus mucosa [19,29]. However, its high expression is a hallmark of many malignant tumors, closely associated with invasion, metastasis, and poor prognosis [20,31]. For instance, in TNBC, EphA2 promotes cell proliferation and bone metastasis [32]; in pancreatic cancer, its overexpression enhances drug delivery efficiency and improves gemcitabine efficacy [33]. Furthermore, high EphA2 expression is closely related to nasopharyngeal carcinoma radiotherapy resistance [34], prostate cancer castration resistance [15], gastric cancer B3GNT3 upregulation [35], and hepatocellular carcinoma CD90-positive cell migration [36]. Overexpression of EFNA5 is also associated with shortened survival in ovarian cancer patients [8]. Therefore, EphA2 is not only an oncogenic signaling hub but also a potential biomarker and therapeutic target.
EphA2 signaling operates through two main modes: ligand-dependent (canonical) and ligand-independent (non-canonical) pathways.
Canonical signaling is triggered by ephrinA1 binding, leading to receptor oligomerization and tyrosine residue autophosphorylation, activating downstream pathways [37]. For example, EphA2 binding to ephrinA1 in vascular smooth muscle cells can inhibit ERK1/2 and AKT activity, thereby suppressing cell migration [29]. Exosomal EphA2 in endothelial cells activates the AMPK signaling pathway via an EphrinA1-dependent pathway, promoting angiogenesis [11].
Non-canonical signaling is activated in the absence of ligand, typically marked by Ser897 phosphorylation [14]. For instance, in prostate cancer, EphA2 S897 phosphorylation promotes castration resistance and invasion [6,15]; in vascular smooth muscle cells, phosphorylation at this site enhances migration [29]. The MK2-RSK signaling axis is considered an important upstream regulator of EphA2 non-canonical activation [14]. Additionally, EphA2 can be cleaved by MT1-MMP into a pro-oncogenic EphA2-NF fragment, serving as a potential tumor biomarker [39]. Thus, canonical signaling often exhibits tumor-suppressive effects, while non-canonical signaling is frequently associated with tumor progression.
The EphA2 signaling network is complex, involving cross-regulation of multiple intracellular pathways, particularly PI3K/AKT, MAPK/ERK, Rho GTPase, and Src family pathways. The mode of activation (ligand-dependent or independent) directly influences the direction and intensity of these downstream signals.
Ligand binding induces EphA2 dimerization and tyrosine autophosphorylation, forming canonical forward signaling [3]. EphA2-ephrinA1 signaling can regulate the cytoskeleton and polarity via the Ezrin-RhoA pathway [1]; control cell adhesion and migration through the Src-FAK-paxillin axis [27]; and inhibit ERK/AKT signaling in vascular smooth muscle cells, preventing pathological proliferation [29]. Furthermore, upon activation, EphA2 can also induce reverse signaling through the ephrin-A ligand, such as exosomal EphA2 activating ERK1/2 or AMPK pathways via EphrinA1, promoting tumor invasion and angiogenesis [11,12]. This indicates that canonical signaling is bidirectional, maintaining cellular homeostasis while also regulating tumor progression in specific contexts.
In the absence of ligand or under specific stress conditions, EphA2 undergoes phosphorylation at Ser897 or Tyr772, forming a non-canonical activation mode [14-16]. Ser897 phosphorylation is mediated by RSK and enhances migration and invasion in prostate cancer, liver cancer, and vascular smooth muscle cells [15,29,36]; chemotherapeutic drugs like cisplatin can also induce S897 phosphorylation via the ERK-RSK-EphA2 pathway, leading to ovarian cancer drug resistance [40]. Phosphorylation at the Y772 site activates Shp2/ERK1/2 signaling, driving nasopharyngeal carcinoma cell proliferation [16]. Additionally, MT1-MMP cleavage, decreased cholesterol, and SNAI1-PIK3R2 interaction can promote EphA2 non-canonical signal activation [10,28,39]. These mechanisms collectively shape EphA2's pro-migration, pro-invasion, and drug resistance properties in tumors.
EphA2 extensively interacts with the PI3K/AKT pathway, often promoting cell survival and migration by upregulating AKT signaling [7,13,35,41,42]. In gastric and breast cancers, EphA2 activates the PI3K/AKT pathway to promote malignant progression [4,35]; whereas in a PTEN-deficient background, Src activation can also upregulate EphA2 expression [45].
MAPK/ERK pathway is another core axis; EphA2 S897 phosphorylation often depends on the ERK-RSK signal [14,46], and its inhibition can restore chemosensitivity [40].
Furthermore, EphA2 regulates the Rho GTPase family to control cytoskeletal remodeling [1,24]; cooperates with Src kinase to regulate focal adhesion dynamics [27,45]; and can drive EMT and stemness maintenance via the Wnt/β-catenin pathway [10,44].
These complex signaling interactions form the biological basis of EphA2 as a "signaling hub" and provide multiple intervention nodes for targeted therapy.
The EphA2 receptor tyrosine kinase, as a key molecule in cellular signal transduction, has broad influence in various pathological processes. This chapter focuses on its functional mechanisms in tumorigenesis and progression, cardiovascular diseases, and inflammatory responses, emphasizing the multiple roles of the EphA2 signaling pathway in disease evolution and treatment.
EphA2 exhibits pro-oncogenic effects in various solid tumors, driving tumor malignancy by regulating cell proliferation, migration, invasion, EMT, angiogenesis, and drug resistance formation. The following sections overview several cancer types where research is most advanced.
In prostate cancer (PCa), high EphA2 expression is closely associated with the progression to castration resistance [6]. Its non-canonical phosphorylation at S897 can activate oncogenic signaling, promoting cell migration, invasion, and dedifferentiation [6,15]. Phosphoproteomic studies reveal that EphA2 activation can induce broad changes in the PI3K/AKT/mTOR and ERK/MAPK pathways, involving cytoskeletal remodeling, motility, and junction regulation [38]. Although EphA2 has dual potential for both pro- and anti-tumor effects, in the CRPC context, its non-canonical signaling clearly favors the pro-oncogenic direction. Adaptor proteins like SHB and afadin further regulate the EphA2-ERK pathway, enhancing cell migration and invasion capabilities [38]. Therefore, blocking EphA2 non-canonical signaling is considered a potential strategy to delay PCa progression.
EphA2 is overexpressed in pancreatic ductal adenocarcinoma (PDAC), representing a potential target for early diagnosis and drug delivery [33]. Studies show that EphA2-targeted gemcitabine conjugate drugs exhibit higher enrichment in tumor tissue, improving efficacy and reducing toxicity [33]. Notably, the matrix metalloproteinase MT1-MMP can cleave the extracellular N-terminus of EphA2, generating the pro-oncogenic fragment EphA2-NF [39]. Serum EphA2-NF levels are significantly elevated in PDAC patients, including early-stage cases, consistent with histological features of EphA2 loss [39]. Therefore, EphA2-NF could serve as a non-invasive biomarker, providing new avenues for early detection and risk assessment of PDAC.
In colorectal cancer (CRC), high EphA2 expression is closely associated with resistance to the EGFR-targeted drug Cetuximab [5]. EphA2 activation can trigger alternative survival signals, diminishing the efficacy of anti-EGFR therapy. The EphA2 inhibitor ALW-II-41-27 combined with Cetuximab can reverse this resistance, inhibiting tumor growth and inducing apoptosis [5]. Clinical analyses also found significantly shorter progression-free survival in patients with high EphA2 expression. On the other hand, CEACAM1-L inhibits CRC liver metastasis by downregulating EphA2 and STAT3 signaling [30], suggesting context-dependent regulation of tumor metastasis by EphA2. In-depth analysis of the interaction between EphA2 and the CEACAM family will help develop new strategies to overcome drug resistance.
High-grade serous ovarian cancer (HGSOC) often relapses due to chemotherapy resistance. Research indicates that EphA2, together with FAK, regulates PI3K/AKT and MAPK/ERK signaling, serving as a key determinant of drug sensitivity [25]. Brigatinib, as a dual inhibitor, can synergize with PARP inhibitors (PARPi) to enhance apoptosis and improve treatment response [25]. Furthermore, platinum-based chemotherapy can induce an ERK1/2-RSK1/2-EphA2-GPRC5A signaling switch, triggering adaptive resistance [40]. RSK inhibitors can prevent EphA2 S897 phosphorylation and restore platinum sensitivity [40]. This finding highlights the central role of EphA2 in chemotherapy resistance formation and provides a new direction for combined targeting of the RSK-EphA2 pathway.
EphA2 is highly expressed in triple-negative breast cancer (TNBC), closely associated with aggressiveness and poor prognosis [31]. EPA combined with an EphA2 inhibitor can significantly enhance apoptosis by disrupting cholesterol homeostasis and inhibiting ABCA1 expression [31]. Moreover, exosomal EphA2 released by highly metastatic breast cancer cells can activate AMPK signaling in endothelial cells, promoting angiogenesis and metastasis [11]. This indicates that EphA2 acts not only as an intra-tumoral signaling molecule but also mediates intercellular communication via exosomes, becoming a key factor in remodeling the tumor microenvironment.
Beyond tumors, EphA2 also plays regulatory roles in cardiovascular and inflammatory diseases. Studies find that EphA2 and ephrinA1 jointly maintain vascular homeostasis in vascular smooth muscle cells (VSMCs), and their ligand binding can inhibit VSMC migration and proliferation [29]. In patients with hyperhomocysteinemia, the EphA2-PI3K/AKT/NF-κB pathway is abnormally activated, leading to impaired endothelial barrier function [43]. In lung inflammation models, EphA2 regulates LPS-induced cytokine release and barrier integrity [17]. Additionally, EphA2 participates in antiviral immune responses in sinus mucosa, influencing the course of chronic rhinitis by regulating epithelial secretion and barrier stability [18,19]. These studies expand the functional scope of EphA2, demonstrating its important role in immunology and vascular biology.
Due to its key role in various diseases, EphA2 has become an emerging focus for targeted therapy. Current drug research primarily concentrates on small molecule inhibitors, antibody-drug conjugates (ADCs), nucleic acid interventions, and nano-delivery systems. Some pipelines under investigation are listed in the table below:
| Drug / Candidate Name | Drug Type | Investigated Indication(s) | Research Institution(s) | Highest R&D Phase |
|---|---|---|---|---|
| Regorafenib | Small Molecule Chemical Drug | Liver Cancer | Hepatocellular Carcinoma | Colorectal Cancer | Gastrointestinal Stromal Tumor | Metastatic Colorectal Cancer, etc. | Bayer AG | Bristol Myers Squibb Co. | Bayer Yakuhin Ltd. | Merck KGaA | Bayer Pharma AG, etc. | Approved; Marketed |
| Dasatinib | Small Molecule Chemical Drug | Accelerated Phase Chronic Myeloid Leukemia | Chronic Phase Philadelphia Chromosome-Positive Chronic Myelogenous Leukemia, etc. | Bristol Myers Squibb Co. | Accord Healthcare SLU | Bristol-Myers Squibb Pharma EEIG , etc. | Approved; Marketed |
| Dasatinib monolauryl sulfate | Small Molecule Chemical Drug | Tumors | Handa Biotech Pharmaceutical Co., Ltd. | New Drug Application |
| Nuzefatide pevedotin (BT5528?) | Peptide-Drug Conjugate (PDC) | Advanced Malignant Solid Tumors | Non-Small Cell Lung Cancer | Ovarian Cancer | Head and Neck Squamous Cell Carcinoma, etc. | Bicycle Therapeutics Plc | Phase 1/2 |
| TP53-EphA-2-CAR-DC vaccine (Chinese PLA General Hospital) | CAR-DC | \- | Zhejiang University | Chinese PLA General Hospital | Phase 1 |
| Dasatinib nanoparticle formulation (Xspray Pharma AB) | Small Molecule Chemical Drug (Nanoparticle) | Philadelphia Chromosome-Positive Chronic Myelogenous Leukemia | Xspray Pharma AB | Phase 1 |
| E-SYNC T Cells (University of California) | CAR-T | EGFR Mutant Glioblastoma | Recurrent Glioblastoma | The University of California, San Francisco | Phase 1 |
| EphA2-targeted CAR-DC (Second Affiliated Hospital, Zhejiang University) | CAR-DC | Metastatic Non-Small Cell Lung Cancer | The Second Affiliated Hospital, School of Medicine, Zhejiang University | Phase 1 |
| P30-EPS vaccine (Duke University) | Synthetic Peptide Vaccine | Therapeutic Vaccine | Glioblastoma Multiforme | Duke University | Phase 1 |
| SC-102 | Peptide-Drug Conjugate (PDC) | Metastatic Solid Tumors | Advanced Malignant Solid Tumors | Solid Tumors by Site | Tianjin StarPeptide Biotechnology Co., Ltd. | Phase 1 |
| siRNA-EphA2-DOPC | siRNA | Ovarian Cancer | Pancreatic Cancer | The University of Texas MD Anderson Cancer Center | Phase 1 |
| MB-108 | Oncolytic Virus | Glioblastoma Multiforme | Gliosarcoma | Recurrent Malignant Glioma | Glioblastoma | The University of Alabama at Birmingham | Nationwide Children's Hospital | Mustang Bio, Inc. | Phase 1 |
| EphA2-targeted CAR-T Cell (Second Affiliated Hospital, Zhejiang University) | CAR-T | Metastatic Non-Small Cell Lung Cancer | The Second Affiliated Hospital, School of Medicine, Zhejiang University | Phase 1 |
(Data as of November 4, 2025, sourced from Synapse)
The EphA2 receptor tyrosine kinase, as a signaling hub, exhibits dual functions in various tumors and inflammatory diseases. CUSABIO provides EphA2 research-related recombinant proteins, antibodies, and ELISA kits to assist your research on the molecular mechanisms of EphA2's bidirectional signaling regulation or the development of EphA2-targeted drugs.
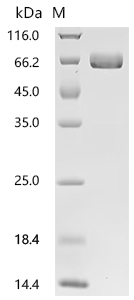
● EphA2 Recombinant Protein
Recombinant Human Ephrin type-A receptor 2 (EPHA2F), partial (Active); CSB-MP007722HUd7
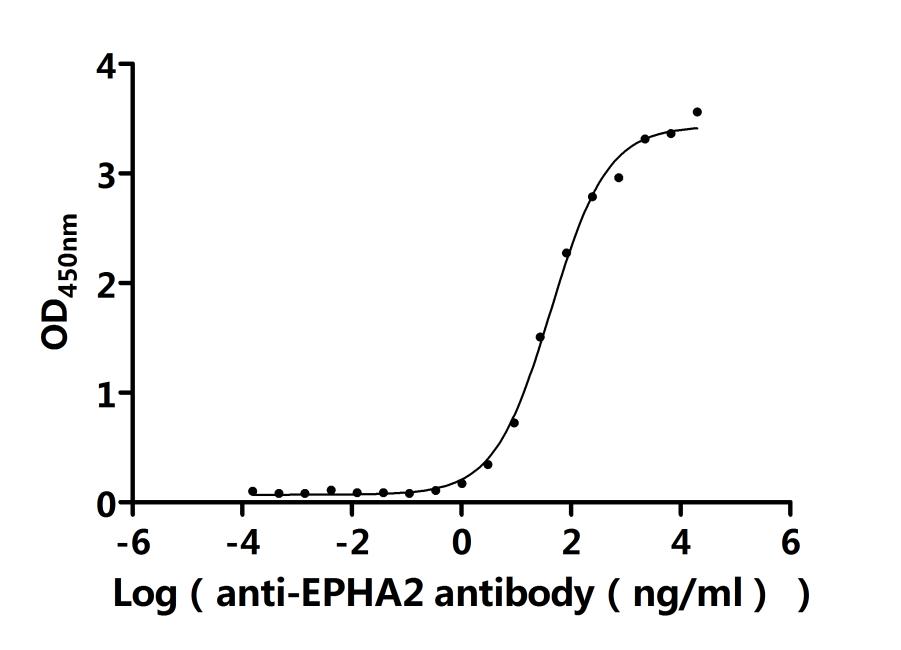
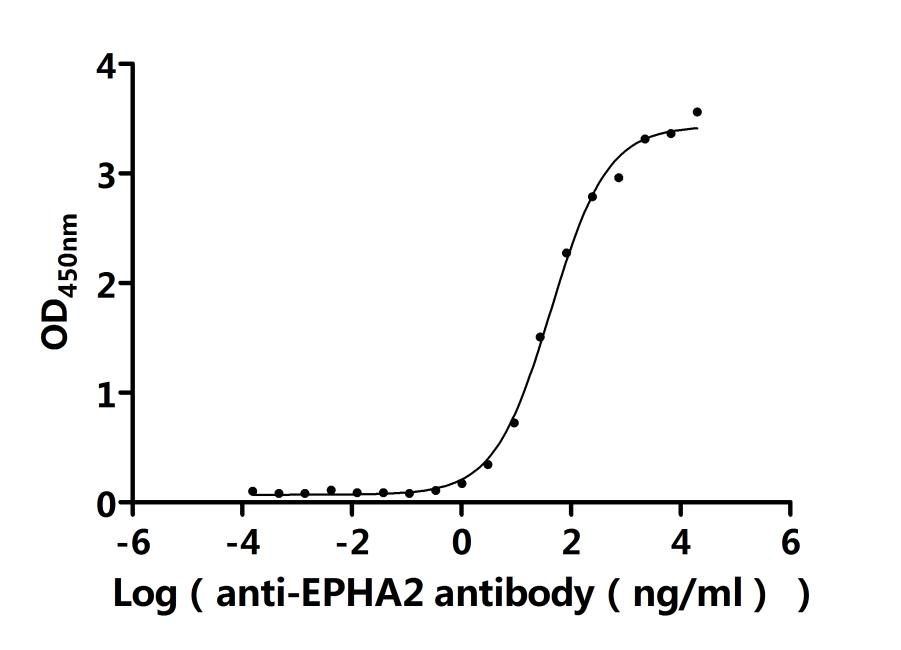
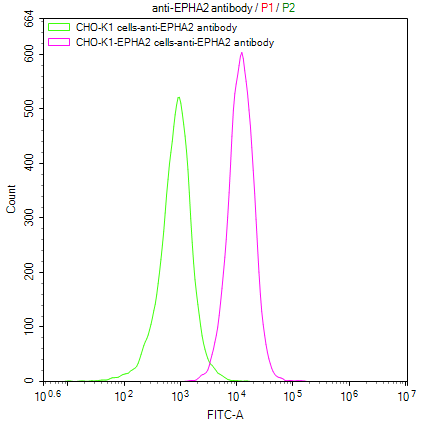
● EphA2 Antibody
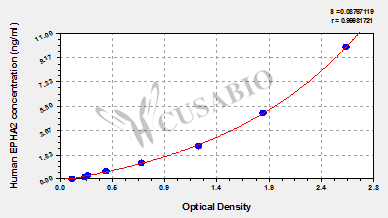
● EphA2 ELISA Kit
● EphA2 Stable Cell Line
References
[1] Yuki Wakayama, K. Miura, H. Sabe, N. Mochizuki.(2011). EphrinA1-EphA2 Signal Induces Compaction and Polarization of Madin-Darby Canine Kidney Cells by Inactivating Ezrin through Negative Regulation of RhoA.
[2] M. Parri, F. Buricchi, M. Taddei, E. Giannoni, G. Raugei, G. Ramponi, P. Chiarugi.(2005). EphrinA1 Repulsive Response Is Regulated by an EphA2 Tyrosine Phosphatase.
[3] Deo R. Singh, Pranjali P Kanvinde, Christopher King, E. Pasquale, K. Hristova.(2018). The EphA2 receptor is activated through induction of distinct, ligand-dependent oligomeric structures.
[4] Xiaoying Huang, Li Na, Qing Han, Qilun Liu, Ligang Wu.(2025). EPHA2 promotes triple-negative breast cancer progression by suppressing pyroptosis via the AKT/PI3K/mTOR pathway.
[5] G. Martini, C. Cardone, P. Vitiello, V. Belli, S. Napolitano, T. Troiani, D. Ciardiello, C. D. Della Corte, F. Morgillo, N. Matrone, V. Sforza, G. Papaccio, V. Desiderio, M. Paul, V. Moreno‐Viedma, N. Normanno, A. Rachiglio, V. Tirino, E. Maiello, T. Latiano, D. Rizzi, G. Signoriello, M. Sibilia, F. Ciardiello, E. Martinelli.(2019). EPHA2 Is a Predictive Biomarker of Resistance and a Potential Therapeutic Target for Improving Antiepidermal Growth Factor Receptor Therapy in Colorectal Cancer.
[6] Ryan Lingerak, A. Petty, Hong Guo, Hebei Lin, Xiaojun Shi, Soyeon Kim, Bingcheng Wang.(2024). Abstract 1680: Roles of EphA2 receptor signaling in prostate cancer development and progression.
[7] Hee-sung Kim, Y. Won, J. Shim, Hyun Ji Kim, B. Kim, H. Hong.(2019). Role of EphA2-PI3K signaling in vasculogenic mimicry induced by cancer-associated fibroblasts in gastric cancer cells.
[8] J. Jukonen, L. Moyano-Galceran, K. Höpfner, E. Pietilä, Laura Lehtinen, K. Huhtinen, E. Gucciardo, J. Hynninen, S. Hietanen, S. Grénman, P. Ojala, O. Carpén, K. Lehti.(2021). Aggressive and recurrent ovarian cancers upregulate ephrinA5, a non-canonical effector of EphA2 signaling duality.
[9] Hayato Kawachi, Tadaaki Yamada, Yuki Katayama, Takayama Koichi.(2025). Abstract 5532: Combinational targeted therapy effects on EphA2 and KRAS G12C inhibitor against KRAS G12C mutated non-small cell lung cancer cells.
[10] H. E, Lei Zhang, Zhenhua Yang, Long Xu, Tao Wang, Junhong Guo, Lang Xia, Juemin Yu, Heyong Wang, Y. She, Junqi Wu, Yue Zhao, Chang Chen, Deping Zhao.(2024). SNAI1 promotes epithelial-mesenchymal transition and maintains cancer stem cell-like properties in thymic epithelial tumors through the PIK3R2/p-EphA2 Axis.
[11] B. Han, He Zhang, Ruinan Tian, Hui Liu, Zhaosong Wang, Zhiyong Wang, Jianfei Tian, Yanfen Cui, Sixin Ren, Xiaoyan Zuo, Ran Tian, R. Niu, Fei Zhang.(2022). Exosomal EPHA2 derived from highly metastatic breast cancer cells promotes angiogenesis by activating the AMPK signaling pathway through Ephrin A1-EPHA2 forward signaling.
[12] Zicong Gao, Xingxing Han, Yuying Zhu, He Zhang, Ran Tian, Zhiyong Wang, Yanfen Cui, Zhaosong Wang, R. Niu, Fei Zhang.(2020). Drug-resistant cancer cell-derived exosomal EphA2 promotes breast cancer metastasis via the EphA2-Ephrin A1 reverse signaling.
[13] Yunyun Wang, Yong Liu, Guo Li, Z. Su, Shuling Ren, Ping-qing Tan, Xin Zhang, Y. Qiu, Yong-quan Tian.(2014). Ephrin type-A receptor 2 regulates sensitivity to paclitaxel in nasopharyngeal carcinoma via the phosphoinositide 3-kinase/Akt signalling pathway.
[14] Fang Zhang, Yue Zhou, Naru Hamada, Akihiro Tanaka, Satoru Yokoyama, Seiji Yano, Kunio Matsumoto, Hiroyuki Mano, Hiroaki Sakurai.(2025). Stress Response Kinase MK2 Induces Non-canonical Activation of EphA2 in EML4-ALK Lung Cancer Cells.
[15] Ryan Lingerak, Bingcheng Wang.(2023). Abstract B018: Loss of androgen receptor-mediated repression leads to EphA2 overexpression that promotes cellular dedifferentiation and castration resistance of prostate cancer through noncanonical signaling.
[16] Yi-Ping Xiang, T. Xiao, Qi-Guang Li, Shan-Shan Lu, Wei Zhu, Yun-Ya Liu, Jie-Ya Qiu, Z. Song, Wei Huang, Hong Yi, Yao-Yun Tang, Zhi-Qiang Xiao.(2020). Y772 phosphorylation of EphA2 is responsible for EphA2-dependent NPC nasopharyngeal carcinoma growth by Shp2/Erk-1/2 signaling pathway.
[17] J. Hong, M. H. Shin, K. Chung, E. Y. Kim, J. Jung, Y. Kang, Y. S. Kim, S. K. Kim, Joon Chang, M. Park.(2015). EphA2 Receptor Signaling Mediates Inflammatory Responses in Lipopolysaccharide-Induced Lung Injury.
[18] J. Shin, M. Han, Jae Hyung Park, Seung Hyeok Lee, Tae Hoon Kim, Sang Hag Lee.(2023). The EphA1 and EphA2 Signaling Modulates the Epithelial Permeability in Human Sinonasal Epithelial Cells and the Rhinovirus Infection Induces Epithelial Barrier Dysfunction via EphA2 Receptor Signaling.
[19] Sang Hag Lee, Sung Hoon Kang, M. Han, Jinsook Kwak, H. Kim, Tae Hoon Lee, Dabin Lee, Tae Hoon Kim.(2021). The Expression of ephrinA1/ephA2 Receptor Increases in Chronic Rhinosinusitis and ephrinA1/ephA2 Signaling Affects Rhinovirus-Induced Innate Immunity in Human Sinonasal Epithelial Cells.
[20] Elisa Fontana, Babar Bashir, Judy S. Wang, R. Aljumaily, J. Machiels, M. Vieito, G. Falchook, Louise Carter, B. D. de Spéville, A. Greystoke, Sang Wun Kim, Nuria Kotecki, A. Spira, I. M. Candilejo, Bristi Basu, H. Prenen, A. Bessudo, Misako Nagasaka, J. Ahnert, Joo-Hwan Park, M. Teo, J. Rotow, Jie Liu, Assunta De Rienzo, Mengyao Li, Adriana Domingo, Hanna Orr, G. Bennett, Rajiv Sharma, M. McKean.(2023). Abstract LB_A17: Trial in progress: First-in-human phase I dose-escalation study of a novel Bicycle toxin conjugate (BT5528) targeting EphA2 in patients with advanced solid tumors.
[21] Alix Tröster, Michael Diprima, Nathalie Jores, D. Kudlinzki, S. Sreeramulu, S. Gande, Verena Linhard, Damian Ludig, Alexander Schug, K. Saxena, Maria Reinecke, S. Heinzlmeir, M. Leisegang, J. Wollenhaupt, Frank Lennartz, M. Weiss, B. Kuster, G. Tosato, H. Schwalbe.(2023). Optimization of the Lead Compound NVP-BHG712 as Colorectal Cancer Inhibitor.
[22] Yige Fu, Drishti Rathod, Ehab M. Abo-Ali, V. Dukhande, Ketan Patel.(2019). EphA2-Receptor Targeted PEGylated Nanoliposomes for the Treatment of BRAFV600E Mutated Parent- and Vemurafenib-Resistant Melanoma.
[23] Jia-Lin Wang, Yu-ling Liu, Y. Li, Wenbing Dai, Zhao-ming Guo, Zhao-hui Wang, Qiang Zhang.(2012). EphA2 targeted doxorubicin stealth liposomes as a therapy system for choroidal neovascularization in rats.
[24] Xia Li, Dan Li, Rong Ma.(2022). ALW-II-41-27, an EphA2 inhibitor, inhibits proliferation, migration and invasion of cervical cancer cells via inhibition of the RhoA/ROCK pathway.
[25] Julie R. Duffield, Xiaonan Hou, Benjamin W. Wilson, Anjali Prasad, Iman K. McKeon-Makki, A. Huehls, Xinyan Wu, Cristina Correia, Melissa C. Larson, F. Couch, A. Oberg, Scott H. Kaufmann, L. Karnitz, Arun Kanakkanthara.(2025). Abstract A024: A PARP-inhibitor-induced early adaptive survival response is tackled through depletion of FRA1 by brigatinib in high-grade serous ovarian carcinoma.
[26] R. Joseph, S. Dasari, Sujanitha Umamaheswaran, L. Mangala, E. Bayraktar, Cristian Rodríguez-Aguayo, Yutuan Wu, N. Nguyen, Reid T Powell, Mary Sobieski, Yuan Liu, Mark S Kim, Sara Corvigno, Katherine I Foster, Pahul Hanjra, Thanh Chung Vu, Mamur A. Chowdhury, P. Amero, Clifford Stephan, G. Lopez-Berestein, S. Westin, Anil K. Sood.(2024). EphA2- and HDAC-Targeted Combination Therapy in Endometrial Cancer.
[27] Zhongwen Chen, D. Oh, K. Biswas, Cheng-han Yu, R. Zaidel-Bar, J. Groves.(2018). Spatially modulated ephrinA1:EphA2 signaling increases local contractility and global focal adhesion dynamics to promote cell motility.
[28] Ryan J Schuck, Alyssa E Ward, A. Sahoo, Jennifer A. Rybak, Robert J Pyron, Thomas N Trybala, Timothy B Simmons, Joshua A. Baccile, Ioannis Sgouralis, Matthias Buck, Rajan Lamichhane, Francisco N. Barrera.(2025). Cholesterol inhibits assembly and oncogenic activation of the EphA2 receptor.
[29] Matthew L. Scott, Alexandra C. Finney, Wayne W Orr.(2023). Abstract 705: Epha2 Ligand-dependent And Ligand-independent Signaling In Vascular Smooth Muscle Cell Proliferation And Migration.
[30] A. Arabzadeh, Kevin McGregor, Valérie Breton, L. Van Der Kraak, U. Akavia, C. Greenwood, N. Beauchemin.(2017). EphA2 signaling is impacted by carcinoembryonic antigen cell adhesion molecule 1-L expression in colorectal cancer liver metastasis in a cell context-dependent manner.
[31] Angie M. Torres-Adorno, H. Vitrac, Y. Qi, L. Tan, K. Levental, Yang-Yi Fan, Peiying Yang, R. Chapkin, Bedrich L Eckhardt, N. Ueno.(2018). Eicosapentaenoic acid in combination with EPHA2 inhibition shows efficacy in preclinical models of triple-negative breast cancer by disrupting cellular cholesterol efflux.
[32] Dominique V. Parker, Verra M. Ngwa, Jin Chen, Julie A. Rhoades.(2024). Abstract 6866: Investigating the role of Epha2 in breast cancer-mediated myeloid cell expansion and function.
[33] B. Quinn, Si Wang, E. Barile, Swadesh K. Das, L. Emdad, D. Sarkar, S. De, Susan Morvaridi Kharagh, J. Stebbins, S. Pandol, P. Fisher, M. Pellecchia.(2016). Therapy of pancreatic cancer via an EphA2 receptor-targeted delivery of gemcitabine.
[34] Jie-Yu Tang, Yun-Xi Peng, Wei Zhu, Jie-Ya Qiu, Wei Huang, Hong Yi, Shan-Shan Lu, Juan Feng, Zheng-Zheng Yu, Di Wu, Qi Wen, Li Yuan, Jinwu Peng, Zhi-Qiang Xiao.(2025). USP5 Binds and Stabilizes EphA2 to Increase Nasopharyngeal Carcinoma Radioresistance.
[35] Hui-Hua Zhou, Jinfeng Zhao, Xiaolin Yang, Jie Liu, Wei Huang.(2022). Study on the Expression of β-1,3-N-acetylglucosaminyltransferase 3 in Gastric Cancer and the Mechanism Promoting Gastric Cancer Progression Based on the Extraction Method of Nanomagnetic Beads.
[36] Nobuhiko Asakura, Naotoshi Nakamura, Atsushi Muroi, Yosui Nojima, T. Yamashita, S. Kaneko, Kazuki Ikeda, N. Koshikawa, Takashi Suzuki.(2021). Expression of Cancer Stem Cell Markers EpCAM and CD90 Is Correlated with Anti- and Pro-Oncogenic EphA2 Signaling in Hepatocellular Carcinoma.
[37] Zhongwen Chen, D. Oh, K. Biswas, R. Zaidel-Bar, J. Groves.(2021). Probing the effect of clustering on EphA2 receptor signaling efficiency by subcellular control of ligand-receptor mobility.
[38] Carolin Offenhäuser, K. Dave, K. Beckett, Fiona M. Smith, Buddhika A. Jayakody, Leanne T. Cooper, Helen Agyei-Yeboah, Jennifer K. McCarron, Yuchen Li, Kate Bastick, F. Al-Ejeh, Jason K. Cullen, M.
Coulthard, Jeffrey J. Gorman, Andrew W. Boyd, B. Day.(2024). EphA2 regulates vascular permeability and prostate cancer metastasis via modulation of cell junction protein phosphorylation.
[39] N. Koshikawa, Shinya Sato, K. Nio, T. Terashima, M. Ueno, Taro Yamashita.(2024). Abstract A059: Serum EphA2 proteolytic fragment is a potent biomarker for diagnosing a very early stage of ductal pancreatic carcinoma.
[40] L. Moyano-Galceran, E. Pietilä, S. Turunen, Sara Corvigno, E. Hjerpe, Daria R Bulanova, U. Joneborg, Twana Alkasalias, Yuichiro Miki, M. Yashiro, A. Chernenko, J. Jukonen, Madhurendra Singh, Hanna Dahlstrand, J. Carlson, K. Lehti.(2020). Adaptive RSK‐EphA2‐GPRC5A signaling switch triggers chemotherapy resistance in ovarian cancer.
[41] Xinxin Gan, Jiatao Hu, Qingyang Pang, Rui Yan, Y. Bao, Ying Liu, Jiaao Song, Zheng Wang, Weihao Sun, Fuzhao Huang, Chen Cai, Linhui Wang.(2024). LDHA‐mediated M2‐type macrophage polarization via tumor‐derived exosomal EPHA2 promotes renal cell carcinoma progression.
[42] Anan Li, Shijiang Wang, Jiangbo Nie, Shining Xiao, Xinsheng Xie, Yu Zhang, Weilai Tong, Geliang Yao, Ning Liu, Fan Dan, Zhiguo Shu, Jiaming Liu, Zhili Liu, Feng Yang.(2024). USP3 promotes osteosarcoma progression via deubiquitinating EPHA2 and activating the PI3K/AKT signaling pathway.
[43] Dan Tian, Q. Qin, Mingfei Li, Xiaoyu Li, Qing Xu, Qian-zhou Lv.(2021). Homocysteine Impairs Endothelial Cell Barrier Function and Angiogenic Potential via the Progranulin/EphA2 Pathway.
[44] Yidan Wang, Zhenting Zhang, Zhengyan Zhu, P. Wang, Jinjuan Zhang, Hui Liu, Jianyu Li.(2022). The significance of EphA2-regulated Wnt/β-catenin signal pathway in promoting the metastasis of HBV-related hepatocellular carcinoma.
[45] Qiong Wang, Xiangyi Kong, Hongming Song, Li Wang, Lingrui Li, Xiaonan Hou, S. Renuse, M. S. Zahari, Ran Cheng, Md Kamrul Hasan Khan, Jidong Wang, Kiran K. Mangalaparthi, Lin Fang, T. Lotan, B. H. Park, S. Weroha, Huaijun Zhou, Akhilesh Pandey, Xinyan Wu.(2025). Proteomic Analysis of PTEN-Deficient Cells Reveals Src-Mediated Upregulation of EphA2 and Therapeutic Potential of Dual Inhibition.
[46] C. Allocca, A. Cirafici, M. Laukkanen, M. Castellone.(2020). Serine 897 Phosphorylation of EPHA2 Is Involved in Signaling of Oncogenic ERK1/2 Drivers in Thyroid Cancer Cells.


Comments
Leave a Comment