Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
Platelet-derived growth factor receptor alpha (PDGFRA) is a receptor tyrosine kinase that plays a critical role in embryonic development, cell proliferation, migration, and differentiation. Its aberrant activation is closely associated with various malignancies, fibrotic diseases, and immune disorders. This article systematically reviews the structural characteristics, signaling mechanisms, roles in multiple diseases, and research progress in targeted drugs for PDGFRA, providing a reference for further basic research and precision therapy.
1. Research Background of PDGFR
2. Structural and Functional Basis of PDGFRA
Platelet-derived growth factor receptors (PDGFRs) belong to the receptor tyrosine kinase (RTK) family and consist of two subtypes: PDGFRA and PDGFRB [1,2]. Among them, PDGFRA primarily mediates signals from ligands such as PDGF-AA, PDGF-AB, and PDGF-CC [3]. During embryonic development, PDGFRA is widely expressed in mesenchymal cells, fibroblasts, and smooth muscle progenitor cells, playing an important role in cell migration, tissue remodeling, and angiogenesis [4].
In pathological states, aberrant PDGFRA signaling is considered a key pathogenic mechanism in various diseases, including tumors, fibrosis, atherosclerosis, and neurological disorders [5,6]. Particularly in gastrointestinal stromal tumors (GIST), PDGFRA gene mutations can lead to conformational changes and constitutive activation of the receptor, representing another major driving factor following KIT mutations [7].
The PDGFRA gene is located on chromosome 4q12 and encodes a transmembrane glycoprotein of approximately 1089 amino acids, containing five immunoglobulin-like extracellular domains, a single transmembrane domain, and an intracellular tyrosine kinase domain [11]. Among these, the third and fourth domains form the ligand-binding region, while the fifth domain is involved in receptor dimerization [12]. Upon receptor activation, its intracellular tyrosine residues are autophosphorylated, providing docking sites for downstream signaling pathways [13].
PDGFRA is highly homologous in structure to PDGFRB but differs in ligand specificity and signal transduction. PDGFRA primarily responds to PDGF-AA, PDGF-AB, and PDGF-CC, whereas PDGFRB is more sensitive to PDGF-BB and PDGF-DD [14].
Furthermore, PDGFRA can undergo cross-phosphorylation with other receptors such as FGFR and EGFR, forming a complex signaling regulatory network [15].
The PDGF family consists of four polypeptide chains (A, B, C, D), which form five hetero/homologous ligands through dimerization: PDGF-AA, PDGF-BB, PDGF-AB, PDGF-CC, and PDGF-DD [16]. These ligands bind to PDGFR subtypes, inducing receptor dimerization and autophosphorylation, thereby activating downstream signaling cascades.
PDGF-AA primarily binds to PDGFRA homodimers (αα), while PDGF-BB can bind to αα, ββ, and αβ complexes [17]. PDGF-CC requires proteolytic cleavage for activation under physiological conditions; its binding to PDGFRA promotes cell migration and angiogenesis [18].
After receptor activation, multiple intracellular tyrosine residues of PDGFRA (e.g., Y572, Y742, Y988, Y1018) are phosphorylated, binding to molecules such as PI3K, PLCγ, Src, and Shp2, thereby triggering multiple signaling pathways [19,20].
PDGFRA plays a crucial role in tissue development and maintenance. During embryogenesis, it is indispensable for the development of the neural crest, heart, lung, and kidneys [21]. Mice lacking PDGFRA exhibit severe developmental defects, including neural tube malformations and impaired angiogenesis [22]. In adult tissues, PDGFRA regulates the proliferation and migration of fibroblasts, smooth muscle cells, and hepatic stellate cells [23]. Additionally, it participates in wound repair and extracellular matrix deposition [24]. Overactivated PDGFRA signaling often leads to excessive cell proliferation and fibrosis, serving as an important pathological basis for various chronic diseases [25].
Upon activation, PDGFRA regulates cell proliferation, differentiation, migration, and survival through several classic signaling pathways, primarily including PI3K/Akt, Ras/MAPK, JAK/STAT, and PLCγ pathways.
The PI3K/Akt pathway is one of the primary survival signals mediated by PDGFRA [26]. After receptor activation, PI3K binds to phosphorylated tyrosine residues (Y742) and is activated, subsequently catalyzing the conversion of PIP2 to PIP3, recruiting Akt to the plasma membrane where it is phosphorylated by PDK1 [27]. Activated Akt promotes the phosphorylation of downstream molecules such as mTOR, GSK3β, and BAD, thereby enhancing cell survival and metabolic activity [28]. In tumor cells, upregulation of the PI3K/Akt signal can inhibit apoptosis and enhance drug resistance [29]. Furthermore, PDGFRA-PI3K/Akt signaling is involved in the differentiation of fibroblasts into myofibroblasts, a key step in fibrosis formation [30].
The Ras/MAPK pathway primarily mediates signals for cell proliferation and migration. Phosphorylated Y988 and Y1018 of PDGFRA recruit the Grb2-SOS complex, activate Ras-GTP, and sequentially initiate the Raf-MEK-ERK cascade reaction [31].
ERK enters the nucleus and promotes the expression of transcription factors such as AP-1, Elk-1, and c-Fos, regulating the transcription of cell cycle proteins (e.g., Cyclin D1) and driving the cell cycle from G1 to S phase [32]. Studies show that constitutive activation of PDGFRA can lead to hyperactivation of the MAPK pathway, causing uncontrolled cell proliferation, particularly in tumors and fibrotic tissues [33].
The JAK/STAT signal is an important pathway through which PDGFRA regulates immune responses and cell differentiation. PDGFRA activation can promote the phosphorylation of JAK1/2, thereby activating STAT1, STAT3, and STAT5 [34]. Activated STAT proteins enter the nucleus and promote the transcription of anti-apoptotic genes (e.g., Bcl-2, Mcl-1) [35]. In glioma and leukemia, upregulation of the PDGFRA-JAK/STAT signal is closely related to tumor cell survival [36].
Simultaneously, this pathway regulates macrophage polarization and cytokine secretion, playing a significant role in chronic inflammation and immune diseases [37].
PDGFRA can also activate the phospholipase Cγ (PLCγ) pathway. Phosphorylation at the Y1021 site of the receptor leads to PLCγ binding, catalyzing the decomposition of PIP2 into IP3 and DAG [38]. IP3 induces the release of Ca²⁺ from the endoplasmic reticulum, while DAG activates PKC, thereby regulating cell migration and contractile functions [39]. This pathway is particularly important in the proliferation and migration of vascular smooth muscle cells; its aberrant activation is closely associated with atherosclerosis and vascular remodeling [40]. Furthermore, Ca²⁺ signaling can provide feedback to regulate PDGFRA phosphorylation levels, forming a dynamic balance mechanism [41].
The signaling network mediated by PDGFRA is highly complex, with cross-regulation existing between different pathways. For example, PI3K/Akt can regulate the intensity of MAPK signaling by inhibiting Raf activation [42]; ERK can provide feedback to regulate PDGFRA phosphorylation, limiting its overactivation [43].
Additionally, PDGFRA can interact with pathways such as TGF-β, EGFR, and VEGFR, forming a multi-level signaling network [44]. In the tumor microenvironment, this crosstalk exacerbates uncontrolled cell growth and drug resistance [45].
Therefore, understanding the dynamic interactions between PDGFRA-related pathways is crucial for precision therapy.
Aberrant activation of PDGFRA is closely associated with various diseases, including malignant tumors, fibrotic diseases, cardiovascular diseases, and neurological disorders. Its mechanisms primarily involve constitutive activation of cell signaling, alterations in the immune microenvironment, and remodeling of the extracellular matrix.
GIST is one of the most extensively studied tumor types regarding PDGFRA. Approximately 10-15% of GISTs harbor PDGFRA mutations, the most common being the D842V mutation in exon 18, which leads to constitutive receptor activation and resistance to imatinib [46]. Avapritinib, as a novel highly selective inhibitor, effectively targets the D842V mutation, significantly improving patient progression-free survival [48].
PDGFRA is frequently amplified or overexpressed in gliomas, particularly in pediatric high-grade gliomas (HGG) [50]. PDGFRA signaling promotes unlimited proliferation of neural glial precursor cells via the PI3K/Akt and STAT3 pathways [51].
In animal models, constitutive activation of PDGFRA can independently drive tumor formation, and combined with p53 loss, it significantly enhances malignancy [52].
PDGFRA is also aberrantly expressed in various solid tumors such as lung adenocarcinoma, colorectal cancer, and breast cancer [55]. Its upregulated signaling is associated with the activation of tumor stromal cells, angiogenesis, and metastasis [56]. For example, in non-small cell lung cancer, PDGF-AA secreted by cancer-associated fibroblasts (CAFs) can activate PDGFRA to promote tumor progression [57]. Inhibiting PDGFRA can reduce tumor cell invasiveness and enhance the response to immunotherapy [58].
PDGFRA signaling is aberrantly activated in fibrosis of multiple organs. It promotes fibroblast proliferation and extracellular matrix deposition, serving as a key driver of fibrogenesis. In liver fibrosis, PDGFRA promotes hepatic stellate cell activation and collagen synthesis via the Akt/mTOR pathway [59]. Inhibiting PDGFRA can significantly alleviate the degree of liver fibrosis [60]. In pulmonary fibrosis models, upregulation of PDGFRA leads to sustained activation of fibroblasts; multi-target TKIs like nintedanib improve disease progression by inhibiting PDGFRA/FGFR/VEGFR signaling [61].
Furthermore, in glomerulosclerosis and myocardial fibrosis, PDGFRA signaling promotes myofibroblast differentiation and enhances ECM deposition, leading to irreversible tissue remodeling [62].
PDGFRA plays a dual role in vascular development and injury repair. Moderate activation promotes vascular smooth muscle proliferation and regeneration, whereas sustained overactivation leads to vascular remodeling and atherosclerosis [47]. Studies show that PDGFRA signaling regulates smooth muscle cell migration via the PLCγ/Ca²⁺ and ERK pathways [49]; its overactivation after arterial injury leads to neointima formation [53]. Inhibiting PDGFRA can reduce abnormal vascular smooth muscle proliferation and improve the risk of restenosis [8]. Therefore, PDGFRA is considered an important potential target for vascular disease intervention.
PDGFRA is also crucial for nervous system development and repair. It is expressed in oligodendrocyte precursor cells (OPCs) and is essential for myelination and regeneration [9]. In neural injury or degenerative diseases, PDGFRA signaling regulates the proliferation of glial cells and axonal regeneration [10]. However, overactivated PDGFRA may lead to abnormal glial cell hyperplasia, associated with the formation of neuroglioma [54]. Therefore, maintaining the balance of PDGFRA signaling is crucial for neural homeostasis.
Currently, drug development targeting PDGFRA exhibits a trend of diversification, covering various types such as small molecule chemicals, monoclonal antibodies, and CAR-T cell therapies. As shown in the table below, in addition to multi-target drugs like ripretinib and avapritinib, which are widely approved for the treatment of gastrointestinal stromal tumors, numerous candidate drugs are in different stages of development, with their indications expanding to include pulmonary arterial hypertension, idiopathic pulmonary fibrosis, soft tissue sarcoma, and other diseases, demonstrating the broad clinical development prospects of this target.
| Medications | Target (Gene Name) | Type of medication | Under investigation indications (disease names) | Institution under Research | Highest Research Stage |
|---|---|---|---|---|---|
| Ripretinib | PDGFRα x c-Kit | Small molecule pharmaceutical | Gastrointestinal stromal tumor | Deciphera Pharmaceuticals, Inc. | Specialised Therapeutics Australia Pty Ltd. | Specialised Therapeutics Asia Pte Ltd. | Zai Lab (Shanghai) Co., Ltd. | Approved for Market |
| Apatinib | PDGFRα x c-Kit | Small molecule pharmaceuticals | Gastrointestinal stromal tumor ;| ;Mastocytosis with myeloid features ;| ;Systemic mastocytosis with aggressive features ;| | Blueprint Medicines (Netherlands) BV | Blueprint Medicines Corp. | KeShi Pharmaceutical | KeShi Pharmaceutical (Suzhou) Co., Ltd. | Approved for Market |
| Olaratumab | PDGFRα | Monoclonal antibody | X-linked ichthyosis | Soft tissue sarcoma | Metastatic soft tissue sarcoma | Sarcoma | Telix Pharmaceuticals Ltd. | Eli Lilly & Co. | Eli Lilly Canada, Inc. | Approved for Market |
| Lenvatinib mesylate | FGFR1 x FGFR2 x FGFR3 x FGFR4 x PDGFRα x RET x VEGFR1 x VEGFR2 x VEGFR3 x c-Kit | Small molecule drugs | Advanced endometrial cancer ;| ;Recurrent endometrial cancer ;| ;Thymoma ;| ;Advanced renal cell carcinoma, etc. | Eisai, Inc. | Merck Sharp & Dohme Corp. | Merck Sharp & Dohme LLC | AiViva BioPharma, Inc. | Eisai GmbH | Eisai Co., Ltd. | Eisai Europe Ltd. | Beijing Tong Ren Tang (Boryung) Co., Ltd. | MSD Korea Co., Ltd. | Jiangsu Shengxian Pharmaceutical Co., Ltd. | Approved for Market |
| Regorafenib | BRAF V600E x CRAF x CSF-1R x DDR2 x EphA2 x FGFR1 x FRK x MAPK11 x PDGFRα x PDGFRβ x RET x Tie-2 x TrkA x VEGFR1 x VEGFR2 x VEGFR3 x c-Kit | Small molecule pharmaceuticals | Liver Cancer | | Hepatocellular Carcinoma | | Colorectal Cancer | | Gastrointestinal Stromal Tumor | | Metastatic Colorectal Cancer, etc. | Merck KGaA | Bayer AG | Bayer HealthCare Pharmaceuticals, Inc. | Bayer Pharma AG | Bristol Myers Squibb Co. | Gustave Roussy, Cancer Campus, Grand Paris | Sun Yat-sen University | Amgen, Inc. | Bayer Yakuhin Ltd. | Bayer HealthCare AG | Approved for Market |
| Pazopanib | FGFR1 x FGFR3 x Flt3L x ITK x LCK x PDGFRα x PDGFRβ x VEGFR1 x VEGFR2 x VEGFR3 x c-Kit | Small molecule drug | Metastatic renal cell carcinoma ;| ;Soft tissue tumors ;| ;Sarcomas ;| ;Renal cell carcinoma ;| ;Soft tissue sarcomas, etc. | Novartis AG | Novartis Pharmaceuticals Corp. | Novartis Pharmaceuticals Australia Pty Ltd. | Novartis Europharm Ltd. | GSK Plc | Novartis Pharma KK | Novartis Pharma Schweiz AG | Approved for Market |
| Seralutinib | CSF-1R x PDGFRα x PDGFRβ x c-Kit | Small molecule pharmaceuticals | Interstitial Lung Disease-Induced Pulmonary Hypertension | Familial Pulmonary Hypertension | Idiopathic Pulmonary Hypertension | Gossamer Bio, Inc. | CHIESI Farmaceutici SpA | Phase 3 Clinical |
| Mesylate Masitinib | LYN x PDGFRα x PDGFRβ x c-Kit | Small molecule pharmaceuticals | Amyotrophic Lateral Sclerosis ;| ;Melanoma ;| ;Metastatic Colorectal Cancer ;etc. | AB Science SA | Clinical Phase 3 |
| Anti-CMV monoclonal antibody (Humabs BioMed SA) | PDGFRα | Monoclonal antibody | Cytomegalovirus Infection | IRB Barcelona | Humabs BioMed SA | Clinical Phase 2 |
| DCC-3009 | PDGFRα x c-Kit | Small molecule drug | Gastrointestinal stromal tumor | Deciphera Pharmaceuticals, Inc. | Phase 1/2 Clinical |
| Ansornitib | DDR1 x DDR2 x PDGFRα x PDGFRβ | Small molecule pharmaceuticals | Idiopathic pulmonary fibrosis | Angion Biomedica Corp. | Clinical Phase 1 |
| ICP-033 | DDR1 x DDR2 x PDGFRα x PDGFRβ x VEGFR2 x VEGFR3 | Small molecule drug | Locally advanced malignant solid tumor | Beijing Nuochengjianhua Pharmaceutical Technology Co., Ltd. | Clinical Phase 1 |
| 89Zr-TLX300-CDx | PDGFRα | Therapeutic Radiopharmaceuticals | Soft Tissue Sarcoma | Telix Pharmaceuticals Ltd. | Clinical Phase 1 |
| IkT-001Pro | ABL x PDGFRα x PDGFRβ x c-Kit | Small Molecule Drug | Chronic Phase Chronic Myeloid Leukemia | Philadelphia Chromosome Positive Chronic Granulocytic Leukemia | Inhibikase Therapeutics, Inc. | Clinical Phase 1 |
| Ubavitinib | PDGFRα x c-Kit | Small molecule pharmaceutical | Late-stage cancer | Advanced malignant solid tumor | Gastrointestinal stromal tumor | Inoperable melanoma | Ningbo Xinyuan Technology Development Co., Ltd. | Ningbo Xinyuan Pharmaceutical Technology Co., Ltd. | Clinical Phase 1 |
| Covalent KIT and PDGFRA Inhibitors (TU Dortmund University) | PDGFRα x c-Kit | Small molecule pharmaceuticals | Gastrointestinal stromal tumor | University of Dortmund | Preclinical |
| PDGFRA Targeted CAR-T (MD Anderson) | PDGFRα | CAR-T | Glioma | University of Texas MD Anderson Cancer Center | Preclinical |
| PDGFR-α inhibitor 6o (China Pharmaceutical University) | PDGFRα | Small molecule drug | Colorectal cancer | China Pharmaceutical University | Preclinical |
| KIT/PDGFRA inhibitor (Tagrisso Biopharmaceuticals) | PDGFRα x c-Kit | Chemical drug | Acute myeloid leukemia | Gastrointestinal stromal tumor | Mastocytosis | Shenzhen Tagiri Biopharmaceutical Co., Ltd. | Preclinical |
| 68Ga-NOTA-CTX004 | PDGFRα | Antibody-Linked Radiopharmaceutical | Diagnostic Radiopharmaceutical | Pancreatic Ductal Adenocarcinoma | Cortalix BV | Preclinical |
| Compound 22 (Shanghai Institute of Materia Medica) | CSF-1R x PDGFRα x SRC family | Small molecule drug | Idiopathic pulmonary fibrosis | Shanghai Institute of Materia Medica, Chinese Academy of Sciences | Preclinical |
| LQFM-064 | PDGFRα x c-Kit x p53 | Small molecule drug | Breast cancer | Federal University of Goiás | Preclinical |
| Compound 4p (Sardar Patel University) | PDGFRα | Small molecule drug | Tumor | Sardar Patel University | Preclinical |
| 6-Hydroxygenistein | PDGFRα x c-Kit | Small molecule pharmaceuticals | Gastrointestinal stromal tumor | Mohammed V University | Preclinical |
(Data as of November 10, 2025, sourced from Synapse)
PDGFRA, as an important receptor tyrosine kinase, plays a central role in various physiological and pathological processes. Its aberrant activation is closely associated with tumors, fibrosis, cardiovascular, and neurological diseases. Cusabio provides PDGFRA recombinant protein, antibodies, and ELISA kits to support your related mechanism research and targeted drug development.
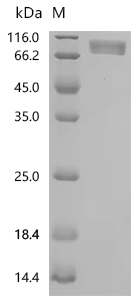
● PDGFRA Recombinant Protein
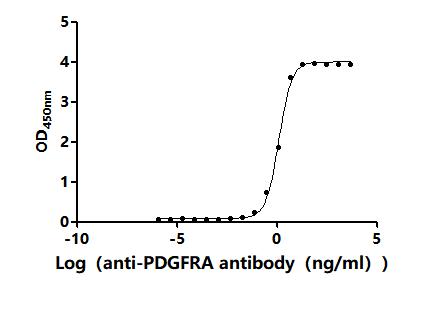
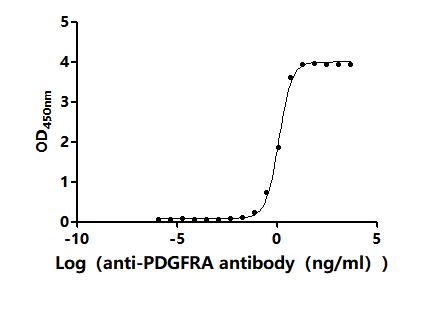
● PDGFRA Antibody
● PDGFRA ELISA Kit
References
[1] Andrew A. Laskin, Nikolai A. Kudryashov, Konstantin G. Skryabin, Eugene V. Korotkov.(2004). Latent periodicity of serine-threonine and tyrosine protein kinases and another protein families.
[2] Rabina Shrestha, Tess McCann, Harini Saravanan, Jaret Lieberth, Prashanna Koirala, Joshua Bloomekatz.(2023). The myocardium utilizes a platelet-derived growth factor receptor alpha (Pdgfra)–phosphoinositide 3-kinase (PI3K) signaling cascade to steer toward the midline during zebrafish heart tube formation.
[3] Katherine A. Fantauzzo, Philippe Soriano.(2014). PI3K-mediated PDGFRα signaling regulates survival and proliferation in skeletal development through p53-dependent intracellular pathways.
[4] Alisa A. Mueller, Cindy T. J. van Velthoven, K. Fukumoto, Tom H. Cheung, T. Rando.(2016). Intronic polyadenylation of PDGFRα in resident stem cells attenuates muscle fibrosis.
[5] Kim van Kuijk, Ian R McCracken, Renée J H A Tillie, Sebastiaan E J Asselberghs, Dlzar A Kheder, Stan Muitjens, Han Jin, Richard S Taylor, Ruud Wichers Schreur, Christoph Kuppe, Ross Dobie, Prakesh Ramachandran, Marion J Gijbels, Lieve Temmerman, Phoebe M Kirkwoord, Joris Luyten, Yanming Li, Heidi Noels, Pieter Goossens, John R Wilson-Kanamori, Leon J Schurgers, Ying H Shen, Barend M E Mees, Erik A L Biessen, Neil C Henderson, Rafael Kramann, Andrew H Baker, Judith C Sluimer.(2023). Human and murine fibroblast single-cell transcriptomics reveals fibroblast clusters are differentially affected by ageing and serum cholesterol.
[6] Elisa Manieri, Guodong Tie, Ermanno Malagola, Davide Seruggia, Shariq Madha, Adrianna Maglieri, Kun Huang, Yuko Fujiwara, Kevin Zhang, Stuart H Orkin, Timothy C Wang, Ruiyang He, Neil McCarthy, Ramesh A Shivdasani.(2023). Role of PDGFRA.
[7] Emilie Guérit, Florence Arts, Guillaume Dachy, Boutaina Boulouadnine, Jean-Baptiste Demoulin.(2021). PDGF receptor mutations in human diseases.
[8] Kentaro Mineji, Jun Watanabe, Eita Uchida, Savneet Kaur, Peng Zhang, Rintaro Hashizume.(2024). DDEL-03. INTRANASAL DELIVERY OF PDGFRA-TARGETED NANOTHERAPEUTICS FOR THE TREATMENT OF PEDIATRIC HIGH-GRADE GLIOMA.
[9] Quirinus J M Voorham, Beatriz Carvalho, Angela J Spiertz, Bart Claes, Sandra Mongera, Nicole C T van Grieken, Heike Grabsch, Martin Kliment, Bjorn Rembacken, Mark A van de Wiel, Philip Quirke, Chris J J Mulder, Diether Lambrechts, Manon van Engeland, Gerrit A Meijer.(2012). Comprehensive mutation analysis in colorectal flat adenomas.
[10] Yiting Deng, Yuanhang He, Juan Xu, Hao He, Manling Zhang, Guang Li.(2025). Cardiac fibroblasts regulate myocardium and coronary vasculature development in the murine heart via the collagen signaling pathway.
[11] T J MacDonald, K M Brown, B LaFleur, K Peterson, C Lawlor, Y Chen, R J Packer, P Cogen, D A Stephan.(2001). Expression profiling of medulloblastoma: PDGFRA and the RAS/MAPK pathway as therapeutic targets for metastatic disease.
[12] Jonathan Paolino, Boris Dimitrov, Beth Apsel Winger, Angelica Sandoval-Perez, Amith Vikram Rangarajan, Nicole Ocasio-Martinez, Harrison K Tsai, Yuting Li, Amanda L Robichaud, Delan Khalid, Charlie Hatton, Riaz Gillani, Petri Polonen, Anthony Dilig, Giacomo Gotti, Julia Kavanagh, Asmani A Adhav, Sean Gow, Jonathan Tsai, Yen Der Li, Benjamin L Ebert, Eliezer M Van Allen, Jacob Bledsoe, Annette S Kim, Sarah K Tasian, Stacy L Cooper, Todd M Cooper, Nobuko Hijiya, Maria Luisa Sulis, Neerav N Shukla, Jeffrey A Magee, Charles G Mullighan, Michael J Burke, Marlise R Luskin, Brenton G Mar, Matthew P Jacobson, Marian H Harris, Kimberly Stegmaier, Andrew E Place, Yana Pikman.(2023). Integration of Genomic Sequencing Drives Therapeutic Targeting of PDGFRA in T-Cell Acute Lymphoblastic Leukemia/Lymphoblastic Lymphoma.
[13] Michele Biscuola, Koen Van de Vijver, María Ángeles Castilla, Laura Romero-Pérez, María Ángeles López-García, Juan Díaz-Martín, Xavier Matias-Guiu, Esther Oliva, José Palacios Calvo.(2013). Oncogene alterations in endometrial carcinosarcomas.
[14] M E Brunkow, D L Nagle, A Bernstein, M Bucan.(1995). A 1.8-Mb YAC contig spanning three members of the receptor tyrosine kinase gene family (Pdgfra, Kit, and Flk1) on mouse chromosome 5.
[15] Carman K M Ip, Patrick K S Ng, Kang Jin Jeong, S H Shao, Zhenlin Ju, P G Leonard, Xu Hua, Christopher P Vellano, Richard Woessner, Nidhi Sahni, Kenneth L Scott, Gordon B Mills.(2018). Neomorphic PDGFRA extracellular domain driver mutations are resistant to PDGFRA targeted therapies.
[16] Qu-Jing Gai, Zhen Fu, Jiang He, Min Mao, Xiao-Xue Yao, Yan Qin, Xi Lan, Lin Zhang, Jing-Ya Miao, Yan-Xia Wang, Jiang Zhu, Fei-Cheng Yang, Hui-Min Lu, Ze-Xuan Yan, Fang-Lin Chen, Yu Shi, Yi-Fang Ping, You-Hong Cui, Xia Zhang, Xindong Liu, Xiao-Hong Yao, Sheng-Qing Lv, Xiu-Wu Bian, Yan Wang.(2022). EPHA2 mediates PDGFA activity and functions together with PDGFRA as prognostic marker and therapeutic target in glioblastoma.
[17] Thomas E Forman, Marcin P Sajek, Eric D Larson, Neelanjan Mukherjee, Katherine A Fantauzzo.(2024). PDGFRα signaling regulates Srsf3 transcript binding to affect PI3K signaling and endosomal trafficking.
[18] M. Riccetti, J. Green, Thomas J Taylor, A. Perl.(2023). Prenatal FGFR2 Signaling via PI3K/AKT Specifies the PDGFRA+ Myofibroblast.
[19] Xuehui Yang, Shivangi Pande, R. Koza, R. Friesel.(2021). Sprouty1 regulates gonadal white adipose tissue growth through a PDGFRα/β-Akt pathway.
[20] Chun-meng Wang, Ying-qiang Shi, Hong Fu, Guang-fa Zhao, Ye Zhou, Chun-yan Du, Yan-wei Ye.(2010). [Oncogenic signaling mechanisms in imatinib-resistant gastrointestinal stromal tumor].
[21] M J Ríos-Moreno, S Jaramillo, M Díaz-Delgado, M Sánchez-León, I Trigo-Sánchez, J Polo Padillo, J Amérigo, R González-Cámpora.(2011). Differential activation of MAPK and PI3K/AKT/mTOR pathways and IGF1R expression in gastrointestinal stromal tumors.
[22] Phillip Zook, H. Pathak, M. Belinsky, Lawrence Gersz, K. Devarajan, Yan Zhou, A. Godwin, M. von Mehren, L. Rink.(2016). Combination of Imatinib Mesylate and AKT Inhibitor Provides Synergistic Effects in Preclinical Study of Gastrointestinal Stromal Tumor.
[23] Martha L Slattery, Lila E Mullany, Lori C Sakoda, Roger K Wolff, John R Stevens, Wade S Samowitz, Jennifer S Herrick.(2018). The PI3K/AKT signaling pathway: Associations of miRNAs with dysregulated gene expression in colorectal cancer.
[24] Zhanghua Wu, Wei Zhao, Zhen Yang, Yue Wang, Yuichi Dai, Liang-an Chen.(2021). Novel Resistance Mechanisms to Osimertinib Analysed by Whole-Exome Sequencing in Non-Small Cell Lung Cancer.
[25] Kiran Kumar Chitluri, Emerson Isaac Arnold.(2025). Integrative genomic analysis identifies DPP4 inhibition as a modulator of FGF17 and PDGFRA downregulation and PI3K/Akt pathway suppression leading to apoptosis.
[26] Olga Martinho, R. Silva-Oliveira, Vera Miranda-Gonçalves, C. Clara, J. R. Almeida, A. Carvalho, J. Barata, R. Reis.(2013). In Vitro and In Vivo Analysis of RTK Inhibitor Efficacy and Identification of Its Novel Targets in Glioblastomas.
[27] Richard J Gilbertson, Jaqueline A Langdon, Andrew Hollander, Roberto Hernan, Twala L Hogg, Amar Gajjar, Christine Fuller, Steven C Clifford.(2006). Mutational analysis of PDGFR-RAS/MAPK pathway activation in childhood medulloblastoma.
[28] Olga Martinho, António Gouveia, Marta Viana-Pereira, Paula Silva, Amadeu Pimenta, Rui Manuel Reis, José Manuel Lopes.(2009). Low frequency of MAP kinase pathway alterations in KIT and PDGFRA wild-type GISTs.
[29] H. Cho, Junfei Zhao, S. Jung, Erik Ladewig, D. Kong, Y. Suh, Yeri Lee, Donggeon Kim, S. Ahn, M. Bordyuh, H. Kang, J. Sa, Y. Seo, S. Kim, D. Lim, Yun-Sik Dho, Jung-Il Lee, H. Seol, J. Choi, W. Park, Chul-Kee Park, R. Rabadán, D. Nam.(2018). Distinct genomic profile and specific targeted drug responses in adult cerebellar glioblastoma.
[30] Herbert M Sauro, Brian Ingalls.(2007). MAPK Cascades as Feedback Amplifiers.
[31] Paul Smolen, Douglas A. Baxter, John H. Byrne.(2008). Bistable MAP Kinase Activity: A Plausible Mechanism Contributing to Maintenance of Late Long-Term Potentiation.
[32] Ruth Nussinov, Chung-Jung Tsai, Carla Mattos.(2013). Pathway drug cocktail: targeting Ras signaling based on structural pathways.
[33] Joanne E Simpson, Noor Gammoh.(2024). Autophagy cooperates with PDGFRA to support oncogenic growth signaling.
[34] Lizhu Liu, Lihong Wu, D. Shan, Bo Han.(2022). Characterization and clinical relevance of PDGFRA pathway copy number variation gains across human cancers.
[35] Tamas Korcsmaros, Illes J. Farkas, Maté S. Szalay, Petra Rovo, David Fazekas, Zoltan Spiro, Csaba Bode, Katalin Lenti, Tibor Vellai, Peter Csermely.(2010). Uniformly curated signaling pathways reveal tissue-specific cross-talks and support drug target discovery.
[36] Cameron W Brennan, Roel G W Verhaak, Aaron McKenna, Benito Campos, Houtan Noushmehr, Sofie R Salama, Siyuan Zheng, Debyani Chakravarty, J Zachary Sanborn, Samuel H Berman, Rameen Beroukhim, Brady Bernard, Chang-Jiun Wu, Giannicola Genovese, Ilya Shmulevich, Jill Barnholtz-Sloan, Lihua Zou, Rahulsimham Vegesna, Sachet A Shukla, Giovanni Ciriello, W K Yung, Wei Zhang, Carrie Sougnez, Tom Mikkelsen, Kenneth Aldape, Darell D Bigner, Erwin G Van Meir, Michael Prados, Andrew Sloan, Keith L Black, Jennifer Eschbacher, Gaetano Finocchiaro, William Friedman, David W Andrews, Abhijit Guha, Mary Iacocca, Brian P O’Neill, Greg Foltz, Jerome Myers, Daniel J Weisenberger, Robert Penny, Raju Kucherlapati, Charles M Perou, D Neil Hayes, Richard Gibbs, Marco Marra, Gordon B Mills, Eric Lander, Paul Spellman, Richard Wilson, Chris Sander, John Weinstein, Matthew Meyerson, Stacey Gabriel, Peter W Laird, David Haussler, Gad Getz, Lynda Chin.(2013). The somatic genomic landscape of glioblastoma.
[37] H. Joensuu.(2023). KIT and PDGFRA Variants and the Survival of Patients with Gastrointestinal Stromal Tumor Treated with Adjuvant Imatinib.
[38] J. Dai, Y. Kong, L. Si, Z. Chi, C. Cui, X. Sheng, L. Mao, Si-ming Li, B. Lian, Ruifeng Yang, Shujing Liu, Xiaowei Xu, Jun Guo.(2013). Large-scale Analysis of PDGFRA Mutations in Melanomas and Evaluation of Their Sensitivity to Tyrosine Kinase Inhibitors Imatinib and Crenolanib.
[39] Tatsuya Kaji, Osamu Yamasaki, Minoru Takata, Masaki Otsuka, Toshihisa Hamada, Shin Morizane, Kenji Asagoe, Hiroyuki Yanai, Yoji Hirai, Hiroshi Umemura, Keiji Iwatsuki.(2017). Comparative study on driver mutations in primary and metastatic melanomas at a single Japanese institute: A clue for intra- and inter-tumor heterogeneity.
[40] Ryan A Denu, Cissimol P. Joseph, Elizabeth Urquiola, Precious S. Byrd, Richard K. Yang, R. Ratan, M. Zarzour, A. Conley, D. Araujo, V. Ravi, E. N. Nassif Haddad, M. Nakazawa, Shreyaskumar R Patel, Wei-Lien Wang, Alexander J. Lazar, N. Somaiah.(2024). Utility of Clinical Next Generation Sequencing Tests in KIT/PDGFRA/SDH Wild-Type Gastrointestinal Stromal Tumors.
[41] Changsong Qi, Fang Pan, Jian Li, Yanyan Li, Jing Gao, Lin Shen.(2018). [Analysis of biological characteristics and prognosis on gastrointestinal stromal tumor with PDGFRA gene mutation].
[42] Michael C Heinrich, Robert G Maki, Christopher L Corless, Cristina R Antonescu, Amy Harlow, Diana Griffith, Ajia Town, Arin McKinley, Wen-Bin Ou, Jonathan A Fletcher, Christopher D M Fletcher, Xin Huang, Darrel P Cohen, Charles M Baum, George D Demetri.(2008). Primary and secondary kinase genotypes correlate with the biological and clinical activity of sunitinib in imatinib-resistant gastrointestinal stromal tumor.
[43] C Serrano, S Bauer, D Gómez-Peregrina, Y-K Kang, R L Jones, P Rutkowski, O Mir, M C Heinrich, W D Tap, K Newberry, A Grassian, H Shi, S Bialick, P Schöffski, M A Pantaleo, M von Mehren, J C Trent, S George.(2023). Circulating tumor DNA analysis of the phase III VOYAGER trial: KIT mutational landscape and outcomes in patients with advanced gastrointestinal stromal tumor treated with avapritinib or regorafenib.
[44] D. Shepherd, T. Miller, D. Forst, P. Jones, V. Nardi, M. Martinez-Lage, A. Stemmer-Rachamimov, R. González, A. Iafrate, Lauren L. Ritterhouse.(2021). Mosaicism for receptor tyrosine kinase activation in a glioblastoma involving both PDGFRA amplification and NTRK2 fusion.
[45] Kohei Kanamori, Y. Yamagata, Y. Honma, Keiichi Date, T. Wada, Tsutomu Hayashi, Sho Otsuki, S. Sekine, T. Yoshikawa, H. Katai, T. Nishida.(2020). Extra-gastrointestinal stromal tumor arising in the lesser omentum with a platelet-derived growth factor receptor alpha (PDGFRA) mutation: a case report and literature review.
[46] Wen Huang, Wei Yuan, Lei Ren, Huaiyu Liang, Xiangyang Du, Xiangfei Sun, Yong Fang, Xiaodong Gao, Min Fu, Yihong Sun, Kuntang Shen, Yingyong Hou.(2022). Clinicopathological and therapeutic analysis of PDGFRA mutated gastrointestinal stromal tumor.
[47] Xuemeng Liu, Yaotian Hu, Z. Xue, Xun Zhang, Xiao-fei Liu, Guowei Liu, Muzi Wen, Anjing Chen, Bin Huang, Xia Li, Ning Yang, Jian Wang.(2023). Valtrate, an iridoid compound in Valeriana, elicits anti-glioblastoma activity through inhibition of the PDGFRA/MEK/ERK signaling pathway.
[48] Michael C Heinrich, Robin L Jones, Margaret von Mehren, Patrick Schöffski, César Serrano, Yoon-Koo Kang, Philippe A Cassier, Olivier Mir, Ferry Eskens, William D Tap, Piotr Rutkowski, Sant P Chawla, Jonathan Trent, Meera Tugnait, Erica K Evans, Tamieka Lauz, Teresa Zhou, Maria Roche, Beni B Wolf, Sebastian Bauer, Suzanne George.(2020). Avapritinib in advanced PDGFRA D842V-mutant gastrointestinal stromal tumour (NAVIGATOR): a multicentre, open-label, phase 1 trial.
[49] S. Abbou, C. Koschmann, C. Kramm, Lindsey M Hoffman, Ashley S. Plant-Fox, M. Abdelbaki, Ashley Bui, M. Casanova, Cornelis M. van Tilburg, Dong-Anh Khuong-Quang, Susan N Chi, Hongliang Shi, I. Bidollari, Janet Hong, P. Swamy, Maria Roche, D. Morgenstern.(2024). TRLS-08. ROVER: A PHASE 1/2 TRIAL IN PROGRESS OF AVAPRITINIB IN PEDIATRIC PATIENTS WITH SOLID TUMORS DEPENDENT ON KIT OR PDGFRA SIGNALING.
[50] R. Olivera-Salazar, Gabriel Salcedo Cabañas, L. Vega-Clemente, David Alonso-Martín, Víctor Manuel Castellano Megías, Peter Volward, D. García-Olmo, M. García-Arranz.(2024). Pilot Study by Liquid Biopsy in Gastrointestinal Stromal Tumors: Analysis of PDGFRA D842V Mutation and Hypermethylation of SEPT9 Presence by Digital Droplet PCR.
[51] Xue Kong, Jun Shi, Dongdong Sun, Lanqing Cheng, Can Wu, Zhiguo Jiang, Yushan Zheng, Wei Wang, Haibo Wu.(2025). A deep-learning model for predicting tyrosine kinase inhibitor response from histology in gastrointestinal stromal tumor.
[52] Pierre Noel.(2012). Eosinophilic myeloid disorders.
[53] C. Koschmann, L. Hoffman, C. Kramm, Ashley S. Plant-Fox, M. Abdelbaki, Ashley Bui, M. Casanova, D. Morgenstern, P. Swamy, Hongliang Shi, Janet Hong, Mikael L Rinne, S. Chi.(2023). TRLS-10. ROVER: A PHASE 1/2 STUDY OF AVAPRITINIB IN PEDIATRIC PATIENTS WITH SOLID TUMORS DEPENDENT ON KIT OR PDGFRA SIGNALING.
[54] Changgong Li, Matt K. Lee, F. Gao, S. Webster, H. Di, J. Duan, Chang-Yo Yang, Navin Bhopal, N. Peinado, G. Pryhuber, Susan M. Smith, Z. Borok, S. Bellusci, P. Minoo.(2019). Secondary crest myofibroblast PDGFRα controls the elastogenesis pathway via a secondary tier of signaling networks during alveologenesis.
[55] Celalettin Ustun, David L DeRemer, Cem Akin.(2011). Tyrosine kinase inhibitors in the treatment of systemic mastocytosis.
[56] Cameron Brennan, Hiroyuki Momota, Dolores Hambardzumyan, Tatsuya Ozawa, Adesh Tandon, Alicia Pedraza, Eric Holland.(2009). Glioblastoma subclasses can be defined by activity among signal transduction pathways and associated genomic alterations.
[57] C. Cobbs, Sabeena Khan, Lisa Matlaf, Sean D McAllister, Alexander Zider, G. Yount, Kenneth Rahlin, L. Harkins, V. Bezrookove, Eric Singer, L. Soroceanu.(2014). HCMV glycoprotein B is expressed in primary glioblastomas and enhances growth and invasiveness via PDGFR-alpha activation.
[58] Soniya Bastola, Marat S Pavlyukov, Neel Sharma, Yasmin Ghochani, Mayu A Nakano, Sree Deepthi Muthukrishnan, Sang Yul Yu, Min Soo Kim, Alireza Sohrabi, Natalia P Biscola, Daisuke Yamashita, Ksenia S Anufrieva, Tatyana F Kovalenko, Grace Jung, Tomas Ganz, Beatrice O’Brien, Riki Kawaguchi, Yue Qin, Stephanie K Seidlits, Alma L Burlingame, Juan A Oses-Prieto, Leif A Havton, Steven A Goldman, Anita B Hjelmeland, Ichiro Nakano, Harley I Kornblum.(2025). Endothelial-secreted Endocan activates PDGFRA and regulates vascularity and spatial phenotype in glioblastoma.
[59] Joanne E Simpson, Morwenna T Muir, Martin Lee, Catherine Naughton, Nick Gilbert, Steven M Pollard, Noor Gammoh.(2024). Autophagy supports PDGFRA-dependent brain tumor development by enhancing oncogenic signaling.
[60] Xiaozhou Yu, Xiao Song, D. Tiek, Runxin Wu, Maya Walker, C. Horbinski, Bo Hu, Shi-Yuan Cheng.(2025). Abstract 6946: Targeting PDGFRa-SHP2 signaling enhances radiotherapy in IDH1 mutant glioma.
[61] Junya Yamaguchi, Fumiharu Ohka, Masafumi Seki, Kazuya Motomura, Shoichi Deguchi, Yoshiki Shiba, Yuka Okumura, Yuji Kibe, Hiroki Shimizu, Sachi Maeda, Yuhei Takido, Ryo Yamamoto, Akihiro Nakamura, Kennosuke Karube, Ryuta Saito.(2024). Dual phenotypes in recurrent astrocytoma, IDH-mutant; coexistence of IDH-mutant and IDH-wildtype components: a case report with genetic and epigenetic analysis.
[62] K. Tateishi, Yohei Miyake, Taishi Nakamura, Hiromichi Iwashita, Takahiro Hayashi, A. Oshima, Hirokuni Honma, Hiroaki Hayashi, Kyoka Sugino, Miyui Kato, K. Satomi, Satoshi Fujii, Takashi Komori, Tetsuya Yamamoto, D. Cahill, H. Wakimoto.(2023). Genetic alterations that deregulate RB and PDGFRA signaling pathways drive tumor progression in IDH2-mutant astrocytoma.


Comments
Leave a Comment