Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
ADAM9, a transmembrane multidomain protein, exhibits both disintegrin and metalloproteinase activities. These functions allow it to modulate a variety of cell-surface proteins via adhesion and ectodomain shedding. Structurally, the ADAM9 protein comprises a signal peptide, a prodomain, a metalloproteinase domain, a disintegrin domain involved in integrin binding, a cysteine-rich region, an EGF-like domain, a transmembrane region, and a short cytoplasmic tail. The prodomain maintains the enzyme in an inactive state; its removal results in the activation of the mature enzyme. The metalloproteinase domain harbors the canonical catalytic activity, which facilitates the cleavage of multiple membrane proteins. The disintegrin domain mediates cell adhesion, while the cytoplasmic tail contains motifs associated with intracellular signaling, allowing ADAM9 to integrate external stimuli with intracellular signaling cascades. The existence of both membrane-bound and secreted ADAM9 isoforms contributes to its functional diversity [1].
Physiologically, ADAM9 is integral to key biological processes such as cell adhesion, migration, protein shedding, and tissue remodeling. It also plays essential roles in inflammatory responses, neural development, and the maintenance of retinal structure. Its metalloproteinase-mediated shedding regulates receptors and adhesion molecules, including Tie-2, VEGFR2, and VCAM-1, thereby influencing angiogenesis, immune cell migration, and intercellular communication. Furthermore, aberrant ADAM9 expression is strongly associated with several pathological conditions, such as degenerative diseases, tissue injury, and various cancers. In the context of tumors, ADAM9 promotes cancer progression, therapeutic resistance, and metastasis through both protease-dependent and -independent mechanisms. Indeed, elevated ADAM9 expression often correlates with a more aggressive clinical phenotype. In summary, the structural diversity and functional complexity of ADAM9 establish it as a pivotal molecule in both maintaining tissue homeostasis and driving disease pathogenesis. Consequently, its distinct molecular characteristics present it as a promising therapeutic target.
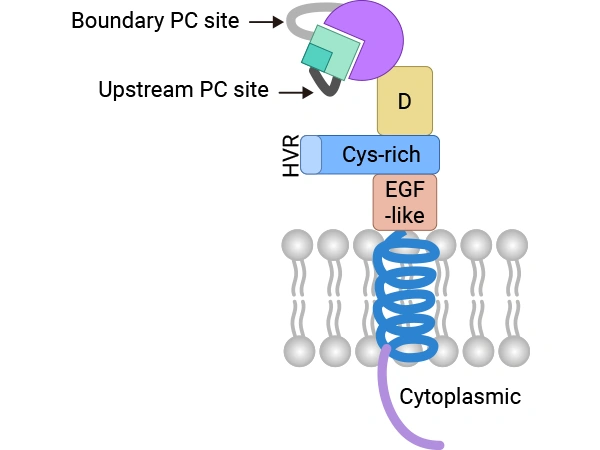
Figure. C-shaped structure of membrane-anchored ADAM9.
doi:10.3390/ijms21207790
As a multifunctional transmembrane protein, ADAM9 is a critical regulator not only in oncology but also in a spectrum of immune and inflammatory diseases, where it influences inflammatory responses, immune cell function, and disease progression. Its unique structural features allow it to participate in intricate signaling networks via both enzymatic and non-enzymatic mechanisms.
In the regulation of the AKT signaling pathway, ADAM9 exerts sophisticated molecular control. Its metalloproteinase domain specifically recognizes and cleaves various membrane-bound precursor proteins, including growth factor receptor ligands. For instance, studies in oral and breast cancer [2] [3] demonstrate that ADAM9-mediated shedding of HB-EGF modulates the activation state of the EGFR family, which in turn activates the downstream PI3K/AKT signaling axis. Critically, ADAM9 can also interact specifically with integrin family members through its disintegrin domain. This interaction alters integrin conformation and function, which subsequently impacts the assembly and activation of the FAK/Src signaling complex [4].
With respect to the NF-κB pathway, ADAM9 demonstrates a more complex, multi-layered regulatory capacity. In addition to proteolytically cleaving TNF family precursor proteins, ADAM9 can directly interact with key regulatory subunits within the IKK complex. This direct interaction potentially modulates the subcellular localization and activation kinetics of the IKK complex [3]. Research also indicates that ADAM9 is involved in the crosstalk between the NF-κB and HIF-1α signaling pathways, a crucial mechanism for tumor adaptation to hypoxic microenvironments [5].
The regulation of tumor angiogenesis by ADAM9 is characterized by notable spatiotemporal specificity. It not only promotes angiogenesis by directly cleaving and releasing VEGF family members but also influences vascular stability by modulating the Angiopoietin-Tie2 signaling system [3] [4]. In pancreatic cancer research, ADAM9 expression levels negatively correlate with vessel maturity, indicating that ADAM9 may foster malignant progression by disrupting vascular normalization [4].
Furthermore, ADAM9 orchestrates the epithelial-mesenchymal transition (EMT) by modulating key signaling nodes [6]. Beyond the traditional TGF-β and Wnt signaling pathways, ADAM9 also regulates the nuclear-cytoplasmic shuttling of the Hippo pathway effectors YAP/TAZ. This regulation may be mediated through the modulation of mechanotransduction, given that ADAM9–integrin interactions can alter the cell’s perception of extracellular matrix stiffness.
In the context of therapeutic resistance, ADAM9 modulates tumor cell stress responses via the establishment of a complex feedback network [3] [7]. During the DNA damage response, ADAM9 not only impacts the activation of the ATM/ATR pathway but also modulates CHK1/2 phosphorylation, thereby regulating cell cycle checkpoint functions. In the regulation of apoptosis, ADAM9 finely tunes mitochondrial outer membrane permeability by influencing the phosphorylation and subcellular localization of Bcl-2 family proteins [7].
ADAM9 can bind to integrins (e.g.,αvβ3,α9β1) to activate the PI3K/Akt and MAPK signaling pathways, thereby promoting neutrophil activation and chemotaxis, which enhances their pro-inflammatory capacity. This process also involves synergistic regulation with CXCR2 signaling, suggesting a role for ADAM9 in facilitating immune cell migration and activation in inflammation-related contexts, such as tumor microenvironments and chronic inflammation [8].
ADAM9 can also subvert immune surveillance by mediating the shedding of various membrane-bound proteins like MICA, TNF-α, and IL-6R. For example, ADAM9-mediated shedding of MICA generates soluble MICA (sMICA), which acts as an immune decoy to inhibit NK cell-mediated tumor clearance, thereby facilitating immune evasion in cancers such as hepatocellular carcinoma and colorectal cancer [9]. Moreover, in chronic hepatitis and non-alcoholic steatohepatitis (NASH), ADAM9 exacerbates fibrosis and carcinogenesis through the regulation of inflammatory cytokines like IL-6.
ADAM9 is a key protein with multiple functions in both physiological and pathological states. Particularly in the field of cancer, it drives tumor progression through various protease-dependent and -independent pathways, making ADAM9 a potential target for drug development. Currently, the predominant type of investigational drugs targeting ADAM9 is Antibody-Drug Conjugates (ADCs), primarily indicated for solid tumors. The following is the pipeline of ADAM9-targeting ADCs in development:
| Drug | Drug Type | Indications | Developers | Highest Phase |
|---|---|---|---|---|
| AEX-6003 | ADC | Tumor | ImmunoGen | Preclinical |
| DB1317 | ADC | Advanced Malignant Solid Tumor,Castration-Resistant Prostate Cancer (CRPC) | Duality Biologics (Suzhou) | Phase I |
| MGC-028 | ADC | Advanced Malignant Solid Tumor,Colorectal Adenocarcinoma, Cholangiocarcinoma, Colorectal Cancer, Non-Small Cell Lung Cancer | MacroGenics | Phase I |
| IMGC-936 | ADC | Advanced Malignant Solid Tumor | MacroGenics, ImmunoGen | Phase I / II |
(Data source: Pharmsnap)
● ADAM9 Recombinant Proteins
(Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
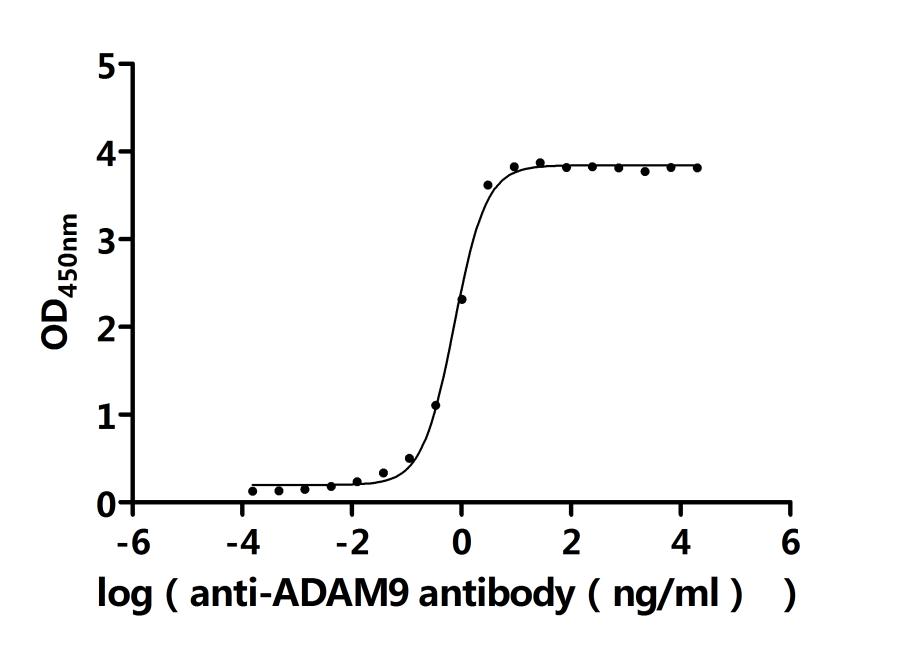
Activity: Measured by its binding ability in a functional ELISA. Immobilized Human ADAM9 at 2μg/ml can bind Anti-ADAM9 recombinant antibody (CSB-RA618774MA1HU). The EC50 is 0.9401-1.088 ng/mL.
Recombinant Macaca fascicularis ADAM9 metallopeptidase domain 9 (ADAM9), Partial,
(CSB-MP7133MOVd7)

(Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
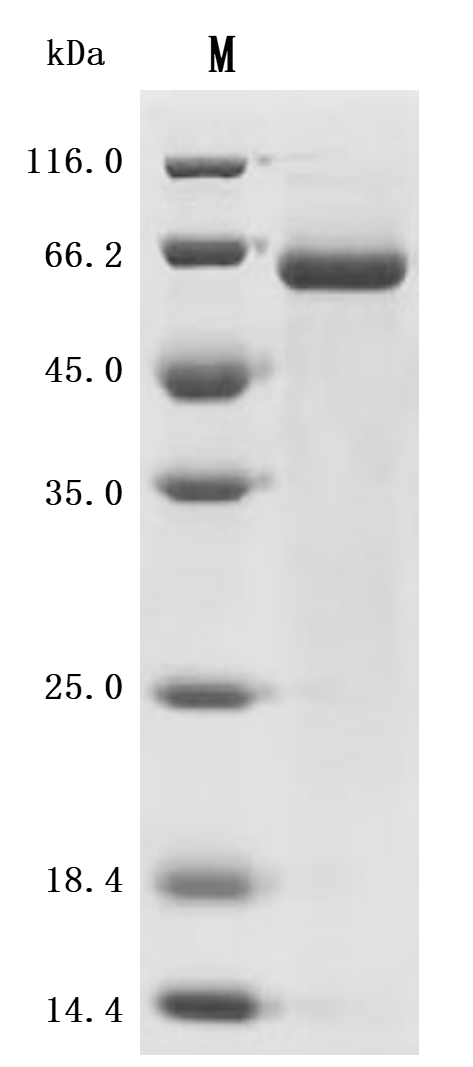
(Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
● ADAM9 Antibodies
ADAM9 Antibody (CSB-PA618774ESR1HU)
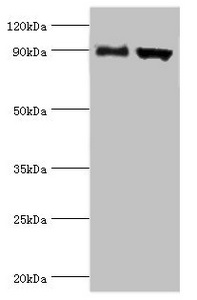
Western blot
All lanes: Disintegrin and metalloproteinase domain-containing protein 9 antibody at 2μg/ml
Lane 1: Mouse liver tissue
Lane 2: Mouse heart tissue
Secondary
Goat polyclonal to rabbit IgG at 1/10000 dilution
Predicted band size: 91,73 kDa
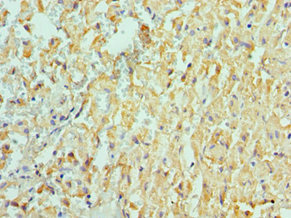
Immunohistochemistry of paraffin-embedded human liver cancer using CSB-PA618774ESR1HU at dilution of 1:100
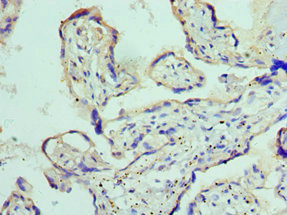
Immunohistochemistry of paraffin-embedded human placenta tissue using CSB-PA618774ESR1HU at dilution of 1:100
● ADAM9 ELISA Kit
References
[1] Chou, C. W., Huang, Y. K., Kuo, T. T., Liu, J. P., & Sher, Y. P. (2020). An Overview of ADAM9: Structure, Activation, and Regulation in Human Diseases. International journal of molecular sciences, 21(20), 7790.
[2] Buranaphatthana, W., Wu, S., Makeudom, A., Sastraruji, T., Supanchart, C., & Krisanaprakornkit, S. (2021). Involvement of the A disintegrin and metalloproteinase 9 in oral cancer cell invasion.. European journal of oral sciences, e12775 .
[3] Zhou, R., Cho, W., , V., Cheuk, W., So, Y., Wong, S., Zhang, M., Li, C., Sun, Y., Zhang, H., Chan, L., & Tian, M. (2020). ADAM9 Mediates Triple-Negative Breast Cancer Progression via AKT/NF-κB Pathway. Frontiers in Medicine, 7.
[4] Oria, V., Lopatta, P., Schmitz, T., Preca, B., Nyström, A., Conrad, C., Bartsch, J., Kulemann, B., Hoeppner, J., Maurer, J., Bronsert, P., & Schilling, O. (2019). ADAM9 contributes to vascular invasion in pancreatic ductal adenocarcinoma. Molecular Oncology, 13, 456 - 479.
[5] Zhu, L., Zhao, Y., Yu, L., He, X., Wang, Y., Jiang, P., Yu, R., Li, W., Dong, B., Wang, X., & Dong, Y. (2021). Overexpression of ADAM9 decreases radiosensitivity of hepatocellular carcinoma cell by activating autophagy. Bioengineered, 12, 5516 - 5528.
[6] Moriwaki, M., Le, T., Sung, S., Jotatsu, Y., Yang, Y., Hirata, Y., Ishii, A., Chiang, Y., Chen, K., Shigemura, K., & Fujisawa, M. (2022). Relevance of A Disintegrin and Metalloproteinase Domain-Containing (ADAM)9 Protein Expression to Bladder Cancer Malignancy. Biomolecules, 12.
[7] Ueno, M., Shiomi, T., Mochizuki, S., Chijiiwa, M., Shimoda, M., Kanai, Y., Kataoka, F., Hirasawa, A., Susumu, N., Aoki, D., & Okada, Y. (2018). ADAM9 is over‐expressed in human ovarian clear cell carcinomas and suppresses cisplatin‐induced cell death. Cancer Science, 109, 471 - 482.
[8] Amêndola, R., Martin, A., Selistre-De-Araújo, H., Paula-Neto, H., Saldanha-Gama, R., & Barja-Fidalgo, C. (2015). ADAM9 disintegrin domain activates human neutrophils through an autocrine circuit involving integrins and CXCR2. Journal of Leukocyte Biology, 97.
[9] Arai, J., Otoyama, Y., Nozawa, H., Kato, N., & Yoshida, H. (2022). The immunological role of ADAMs in the field of gastroenterological chronic inflammatory diseases and cancers: a review. Oncogene, 42, 549 - 558.


Comments
Leave a Comment