Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
| Code | CSB-YP355623MO |
| Abbreviation | Recombinant Mouse Ins1 protein |
| MSDS | |
| Size | $368 |
| Order now | |
| Image |
|
| Have Questions? | Leave a Message or Start an on-line Chat |
There are currently no reviews for this product.
Has CSB-YP355623MO been tested in cell culture?
I have some problems in the development of the immunoassay to detect mouse insulin. Our antibodies are produced against recombinant human insulin, but they also react with mouse insulin (affinity tested). However, once I perform the immunoassay I could not see any specific signal, all signal is like background noise. I¡¯m not sure if either insulin or one of the antibodies is not working. However, I have some questions regarding recombinant mouse insulin that I hope you could answer,
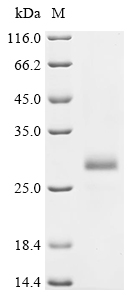
Insulin purity was assessed by SDS-PAGE, addressing 85 % of purity, I don¡¯t understand why stained gel only shows one single band, theoretically there are other proteins/peptides, which could show more other bands in SDS-PAGE, right? . In this sense, if insulin expected band should be at 13 kDa (in you datasheet), why stained gel shows a single band at 21 kDa. I guess that this insulin is suitable as a standard/calibrator for immunoassays development, right? (Of course, always depending on the antibodies used).

