Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
MYL9, the regulatory light chain of myosin, is essential for cell movement by activating myosin motility. It has been shown that MYL9 promotes cancer cell migration and is implicated in tumor development and progression. Interestingly, MYL9 has also been found to interact with CD69, an early activation marker of lymphocytes. This interaction enables the recruitment of T cells to the inflamed lung, which is significant in understanding immune responses [1].
Moreover, the MYL9-CD69 system depletes effector T cells in the tumor microenvironment, weakening the anti-tumor immune response. By disrupting the interaction between MYL9 and CD69, researchers found that it can potentially boost the anti-tumor immune response, make MYL9 an attractive target for cancer immunotherapy [2]. Considering its association with the myosin motors family, MYL9 holds promise as a novel drug target for cancer immunotherapy!
Myosins are conserved proteins that serve as actin-dependent molecular motors, playing important roles in the cytoskeleton. They make up 15%-25% of total body proteins and are mainly found in smooth muscle. The diverse human myosins family consists of 12 classes (I-XII) with 40 identified myosin genes. Myosin's molecular structure includes heavy chains (MHC) and light chains (MLC), which are further classified into basic MLC (MLC I) and regulatory MLC (MLC II). Functionally, myosin motors have emerged as powerful players that drive cytoplasmic streaming, actin organization, and cell expansion (Figure 1) [3-6].
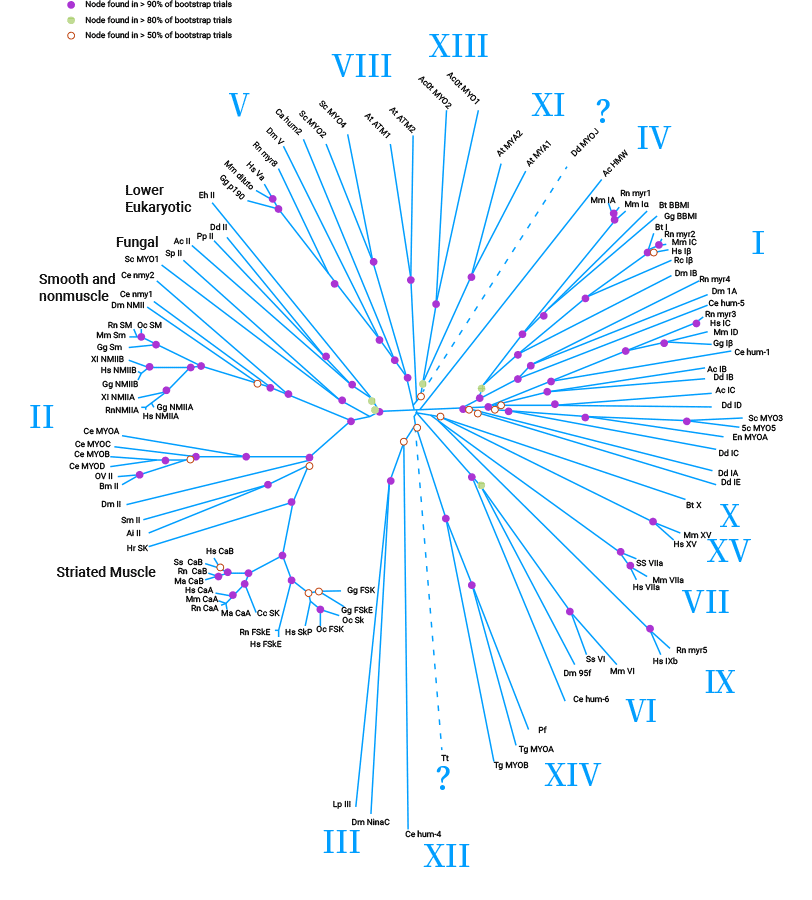
Figure 1. Myosin motors superfamily [3]
MYL9, also known as MLC2, MRLC1, or MLC-2C, is an important component of myosin. The human MYL9 gene is located on chromosome 20q11.23 and consists of 8621 bases, with 8 introns and 17 exons. It encodes the 20KD myosin regulatory light chain. Myosin is composed of the head, neck, and tail regions. The head has ATP-binding and actin-binding sites for ATP hydrolysis, energy release, and actin binding. The neck contains an alpha helix entwined by two regulatory light chains that bind to myosin light chains and calmodulin. The tail forms a coiled helical structure with hydroxyl groups (Figure 2) [7].
MYL9 is expressed extensively in both normal and tumor tissues, including muscle tissue, visceral tissue, lung cancer, and prostate cancer, etc. Numerous studies have demonstrated that MYL9, as a crucial member of the myosin family, participates in various organismal functions regulated by diverse factors. These functions encompass muscle movement, cell migration, endocytosis, mitosis, signal transduction, and notably, the migration and proliferation of tumor cells [8-11].
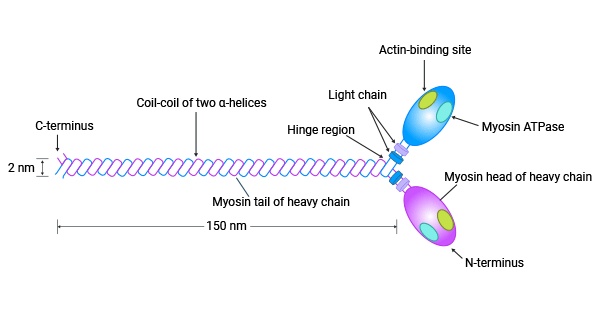
Figure 2. MYL9 is a power player in the myosins [7]
MYL9 exerts regulatory functions through two major systems: the Rho-kinase (ROCK) system and the myosin light chain kinase (MLCK) system. Additionally, several other important molecules such as calmodulin-dependent protein kinase II, ILK, PKA, ZIPK, PKC, and the inhibitory molecule CP17 of myosin light chain phosphatase MLCP are also involved [12-14].
As shown in Figure 3, the MLCK and ROCK systems are activated by Ca2+, which leads to an increase in intracellular Ca2+ levels. This activation triggers the activation of myosin light chain myosin. Specifically, ROCK acts as a substrate for myosin light chain, facilitating the phosphorylation and dephosphorylation of the MYL9 gene. Besides, ROCK promotes the elevation of intracellular Ca2+ concentration, which activates MLCK [12].
Phosphorylating MYL9 improves the interaction between myosin and actin, as well as enhances ATPase activity in the myosin light chain head. This leads to cytoskeletal remodeling and boosts cellular capabilities in proliferation, differentiation, adhesion, and migration. The RhoA inhibitory factor effectively hinders the downstream effector protein Rock, thereby inhibiting MYL9 expression and phosphorylation. Consequently, it suppresses actin formation, tumor invasion, metastasis, and tumor cell growth [12].
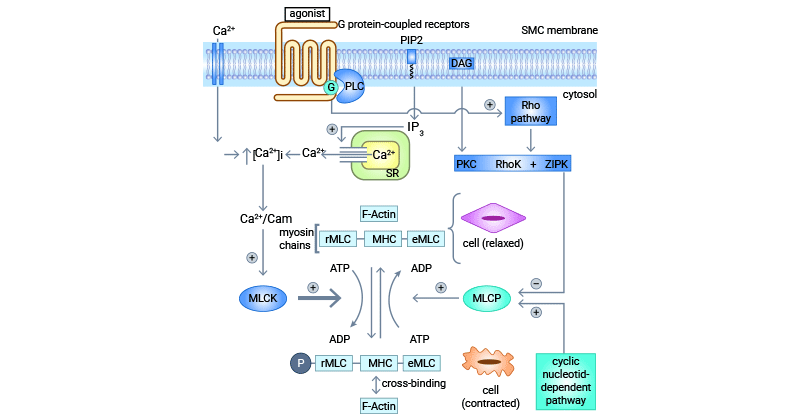
Figure 3. MYL9 plays a regulatory role through MLCK and ROCK systems [12]
Numerous studies have shown that MYL9 regulates ATPase activity and myosin contraction through phosphorylation and dephosphorylation. Abnormal expression of MYL9, which is associated with cell motility. Various investigations have reported the functions of MYL9 in cell motility is linked to tumor pathogenesis, with effects ranging from tumor suppression to tumor promotion [15-17].
MYL9 is increased in various tumors and contributes to tumor invasion in breast, liver, and glioblastoma cancers. It enhances tumor cell motility in breast and liver cancers [18-19]. In glioblastoma, higher MYL9 expression and phosphorylation levels are associated with poor prognosis and recurrent cases [8]. Pancreatic ductal adenocarcinoma and ovarian epithelial tumors also show increased MYL9 expression. Clinical analysis suggests that MYL9 could be an independent prognostic factor in these tumors [14, 20].
Similarly, MYL9 is significantly upregulated in esophageal squamous cell carcinoma and is associated with poorer overall survival and recurrence-free survival in patients with high MYL9 expression [21]. It also contributes to the progression of osteosarcoma [22]. These findings suggest that MYL9 enhances tumor cell motility, potentially affecting tumor growth and metastasis. Moreover, MYL9 shows promise as a prognostic marker and therapeutic target for various types of tumors.
In contrast, in prostate tissue, MYL9 protein expression was decreased and correlated with age, pathological stage, metastasis, and PSA levels [23]. Similarly, downregulation of MYL9 expression was found in bladder and gastric cancers [24-25]. However, cellular experiments revealed that MYL9 deficiency reduced proliferation and increased apoptosis in gastric cancer cells [15]. In human colorectal cancer, both MYL9 expression and phosphorylation levels were reduced [26]. Upregulating MYL9 inhibited tumor cell proliferation, invasion, and migration while promoting apoptosis in colon cancer stem cells [27-28].
Low MYL9 expression may be associated with non-small cell lung cancer development and metastasis [29-30]. In breast cancer, increased MYL9 reduces cancer cell motility [15]. However, conflicting reports suggest that MYL9 expression at the molecular level may enhance breast cancer cell migration [15, 31]. Thus, the role of MYL9 in malignancy remains controversial, and its involvement in tumors is multifaceted. This complexity underscores the potential value of MYL9 as a valuable marker for therapeutic targeting and prognostic assessment.
Apart from its role in tumors, MYL9 is also implicated in cardiovascular, inflammatory, and neurological diseases [32-35]. Studies on atherosclerosis models have shown that MYL9 expression is significantly downregulated as plaque increases. MYL9 is influenced by angiotensin II (Angiotensin II/AGT) and cytokines such as PDGF-BB. Angiotensin II and PDGF-BB are crucial cytokines that promote smooth muscle cell proliferation and migration during vascular injury. Therefore, MYL9 may have an important role in the development of atherosclerosis [36-37].
Successive reports suggested that MYL9 serves as a new ligand for CD69, crucially involved in regulating immune responses [2, 38-41]. Blocking the interaction between MYL9 and CD69 has demonstrated improvement in allergic respiratory inflammation, such as asthma [2]. During inflammation, vascular damage, and platelet activation, MYL9 network structures and coagulation factors are produced. This leads to the binding of MYL9 to CD69+T cells, resulting in the aggregation of immune cells at the sites of inflammation. Consequently, effector cytokines and chemokines are generated, effectively triggering the activation of the immune response (Figure 4) [41].
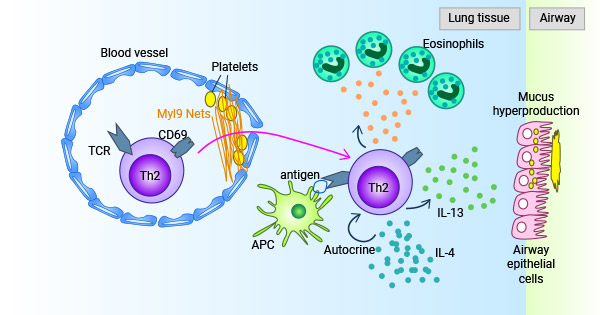
Figure 4. The MYL9-CD69 system plays a role in immune response [42]
MYL9 is overexpressed in various cancers and associated with poor prognosis. It may act as a ligand for CD69, promoting the retention of cytotoxic T lymphocytes (CTLs) in the tumor microenvironment. This chronic stimulation by tumor antigens leads to T-cell failure. Therefore, targeting the MYL9/CD69 interaction shows promise for cancer immunotherapy due to the important role of antigen-specific CTLs in defending against tumors.
Currently, the clinical experience with MYL9 in cancer treatment is limited. However, mounting evidence supports its potential as a therapeutic target for various tumor treatments. Consequently, MYL9 holds significant clinical application as a molecular marker and potential target for early diagnosis, prognosis prediction, and targeted therapy of tumors.
To fully support researchers and pharmaceutical companies in their research on MYL9 in tumors or other diseases, CUSABIO presents MYL9 protein to support your research on the mechanism of MYL9 or its potential clinical value (click for the full list of MYL9 products: MYL9 Proteins; MYL9 antibodies).
Recombinant Human Myosin regulatory light polypeptide 9(MYL9) (Active) (Code: CSB-YP015318HU)
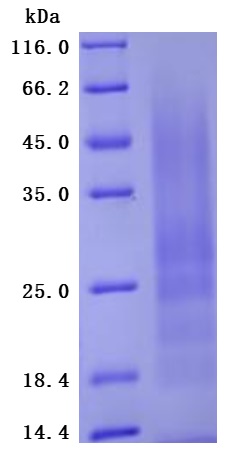
The greater than 95% as determined by SDS-PAGE.(Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
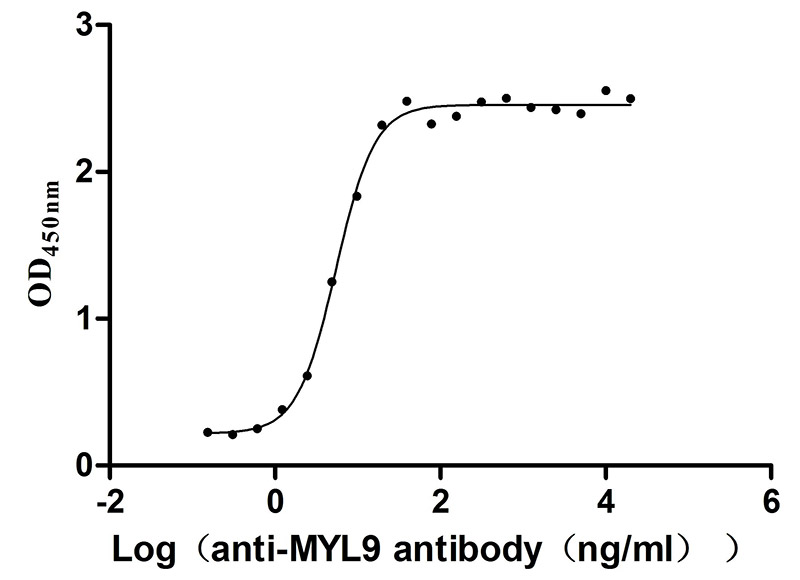
Immobilized Human MYL9 at 2μg/mL can bind Anti-MYL9 recombinant antibody (CSB-RA015318MA1HU), the EC50 is 4.628-6.430 ng/mL.
References
[1] Iwamura, Chiaki, et al. "Elevated Myl9 reflects the Myl9-containing microthrombi in SARS-CoV-2-induced lung exudative vasculitis and predicts COVID-19 severity." Proceedings of the National Academy of Sciences 119.33 (2022): e2203437119.
[2] Kimura, Motoko Y., et al. "A new therapeutic target: the CD69-Myl9 system in immune responses." Seminars in immunopathology. vol. 41. Springer Berlin Heidelberg, 2019.
[3] Sellers, James R. "Myosins: a diverse superfamily." Biochimica et Biophysica Acta (BBA)-Molecular Cell Research 1496.1 (2000): 3-22.
[4] Fitz, Gillian N., et al. "Protrusion growth driven by myosin-generated force." Developmental Cell 58.1 (2023): 18-33.
[5] Hartman, M. Amanda, and James A. Spudich. "The myosin superfamily at a glance. "Journal of cell science 125.7 (2012): 1627-1632.
[6] Lewis, Christopher TA, and Julien Ochala. "Myosin heavy chain as a novel key modulator of striated muscle resting state. "Physiology 38.1 (2023): 3-9.
[7] Bhanu P. Jena. "Myosin: Cellular Molecular Motor" Cellular Nanomachines (2020): 79-89.
[8] Kruthika, Banavathy S., et al. "Expression pattern and prognostic significance of myosin light chain 9 (MYL9): a novel biomarker in glioblastoma." Journal of Clinical Pathology 72.10 (2019): 677-681.
[9] Shehadeh, Lina A., et al. "Dynamic regulation of vascular myosin light chain (MYL9) with injury and aging." PLoS One 6.10 (2011): e25855.
[10] Zhu, Kongxi, et al. "Long non-coding RNA MBNL1-AS1 regulates proliferation, migration, and invasion of cancer stem cells in colon cancer by interacting with MYL9 via sponging microRNA-412-3p." Clinics and research in hepatology and gastroenterology 44.1 (2020): 101-114.
[11] Jalagadugula, Gauthami, et al. "Regulation of platelet myosin light chain (MYL9) by RUNX1: implications for thrombocytopenia and platelet dysfunction in RUNX1 haplodeficiency." Blood, The Journal of the American Society of Hematology 116.26 (2010): 6037-6045.
[12] Moreno, Carolina Araujo, et al. "Homozygous deletion in MYL9 expands the molecular basis of megacystis-microcolon- intestinal hypoperistalsis syndrome." European Journal of Human Genetics 26.5 (2018): 669-675.
[13] Licht, Alexander H., et al. "Junb regulates arterial contraction capacity, cellular contractility, and motility via its target Myl9 in mice." The Journal of clinical investigation 120.7 (2010): 2307-2318.
[14] Matsushita, Katsunori, et al. "Clinicopathological significance of MYL9 expression in pancreatic ductal adenocarcinoma." Cancer Reports 5.10 (2022 ): e1582.
[15] Lv, Minghe, Lumeng Luo, and Xue Chen. "The landscape of prognostic and immunological role of myosin light chain 9 (MYL9) in human tumors." Immunity. Inflammation and Disease 10.2 (2022): 241-254.
[16] Gilles, Laure, et al. "MAL/SRF complex is involved in platelet formation and megakaryocyte migration by regulating MYL9 (MLC2) and MMP9." Blood, The Journal of the American Society of Hematology 114.19 (2009): 4221-4232.
[17] Tan, Xiang, and Mingwu Chen. "MYLK and MYL9 expression in non-small cell lung cancer identified by bioinformatics analysis of public expression data." Tumor Biology 35 (2014): 12189-12200.
[18] Luo, Xue-Gang, et al. "Histone methyltransferase SMYD3 promotes MRTF-A-mediated transactivation of MYL9 and migration of MCF-7 breast cancer cells." Cancer letters 344.1 (2014): 129-137.
[19] Hirasawa, Yuichi, et al. "Methylation status of genes upregulated by demethylating agent 5-aza-2′-deoxycytidine in hepatocellular carcinoma." Oncology 71.1-2 (2007): 77-85.
[20] Deng, Yuao, et al. "High expression of MYL9 indicates poor clinical prognosis of epithelial ovarian cancer." Recent Patents on Anti-Cancer Drug Discovery 16.4 (2021): 533-539.
[21] Wang, Jian-Hua, et al. "Expression and prognostic significance of MYL9 in esophageal squamous cell carcinoma." ploS one 12.4 (2017): e0175280.
[22] Zhao, S., W. Xiong, and K. Xu. "MiR-663a, regulated by lncRNA GAS5, contributes to osteosarcoma development through targeting MYL9. "Human & Experimental Toxicology 39.12 (2020): 1607-1618.
[23] Huang, Ya-qiang, et al. "Decreased expression of myosin light chain MYL9 in stroma predicts malignant progression and poor biochemical recurrence- free survival in prostate cancer." medical oncology 31 (2014): 1-9.
[24] Huang, Chu-Han, et al. "MYL9 deficiency is neonatal lethal in mice due to abnormalities in the lung and the muscularis propria of the bladder and intestine ." Plos one 17.7 (2022): e0270820.
[25] Dong, Ningxin, et al. "Identification and validation of critical genes with prognostic value in gastric cancer." Frontiers in Cell and Developmental Biology 10 (2022): 1072062.
[26] Qiu, Xiao, et al. "Weighted gene co-expression network analysis identified MYL9 and CNN1 are associated with recurrence in colorectal cancer." journal of Cancer 11.8 (2020): 2348.
[27] Zhou, Rui, et al. "PRPF19 facilitates colorectal cancer liver metastasis through activation of the Src-YAP1 pathway via K63-linked ubiquitination of MYL9." Cell Death & Disease 14.4 (2023): 258.
[28] Feng, Min, et al. "Myosin light chain 9 promotes the proliferation, invasion, migration and angiogenesis of colorectal cancer cells by binding to Yes- associated protein 1 and regulating Hippo signaling." Bioengineered 13.1 (2022): 96-106.
[29] Sheng, Meiling, Zhaohui Dong, and Yanping Xie. "Identification of tumor-educated platelet biomarkers of non-small-cell lung cancer. "OncoTargets and therapy (2018): 8143-8151.
[30] Tan, Xiang, and Mingwu Chen. "MYLK and MYL9 expression in non-small cell lung cancer identified by bioinformatics analysis of public expression data." Tumor Biology 35 (2014): 12189-12200.
[31] Zhang, Chunling, et al. "Myocardin-related transcription factor A is up-regulated by 17β-estradiol and promotes migration of MCF-7 breast cancer cells via transactivation of MYL9 and CYR61." Acta Biochim Biophys Sin 45.11 (2013): 921-927.
[32] Kandler, Justin L., et al. "Compound heterozygous loss of function variants in MYL9 in a child with megacystis-microcolon intestinal hypoperistalsis syndrome." Molecular Genetics & Genomic Medicine 8.11 (2020): e1516.
[33] Kobayashi, Hironobu, et al. "Increased Myosin light chain 9 expression during Kawasaki disease vasculitis." Frontiers in Immunology 13 (2023). 1036672.
[34] Xiong, Yao, et al. "Targeting MRTF/SRF in CAP2-dependent dilated cardiomyopathy delays disease onset." jci insight 4.6 (2019).
[35] Sun, L., et al. "Decreased platelet expression of myosin regulatory light chain polypeptide (MYL9) and other genes with platelet dysfunction and CBFA2/ RUNX1 mutation: insights from platelet expression profiling." Journal of Thrombosis and Haemostasis 5.1 (2007): 146-154.
[36] Wen, Dingke, et al. "Single-cell RNA sequencing reveals the pathogenic relevance of intracranial atherosclerosis in blood blister-like aneurysms." Frontiers in Immunology 13 (2022): 927125.
[37] Hwang, Ki-Chul. "The Role of MicroRNAs in Vascular Diseases; Smooth Muscle Cell Differentiation and De-Differentiation. "Korean Circulation Journal 44.4 (2014): 218-219.
[38] Hayashizaki, Koji, et al. "Myosin light chains 9 and 12 are functional ligands for CD69 that regulate airway inflammation." science immunology 1.3 (2016 ): eaaf9154-eaaf9154.
[39] Nakayama, Toshinori, et al. "CD4+ T cells in inflammatory diseases: pathogenic T-helper cells and the CD69-Myl9 system." International Immunology 33.12 (2021): 699-704.
[40] Yokoyama, Masaya, et al. "Myosin light chain 9/12 regulates the pathogenesis of inflammatory bowel disease." Frontiers in Immunology 11 (2021): 594297.
[41] Onodera, Atsushi, Kota Kokubo, and Toshinori Nakayama. "Epigenetic and transcriptional regulation in the induction, maintenance, heterogeneity and recall-response of effector and memory Th2 cells." Frontiers in Immunology 9 (2018): 2929.


Comments
Leave a Comment