
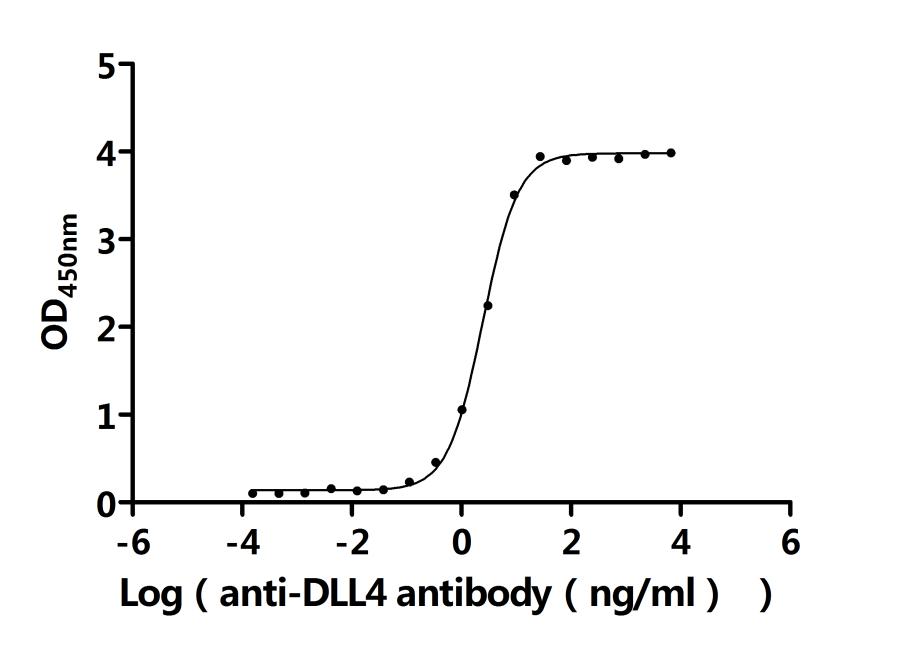
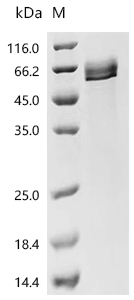
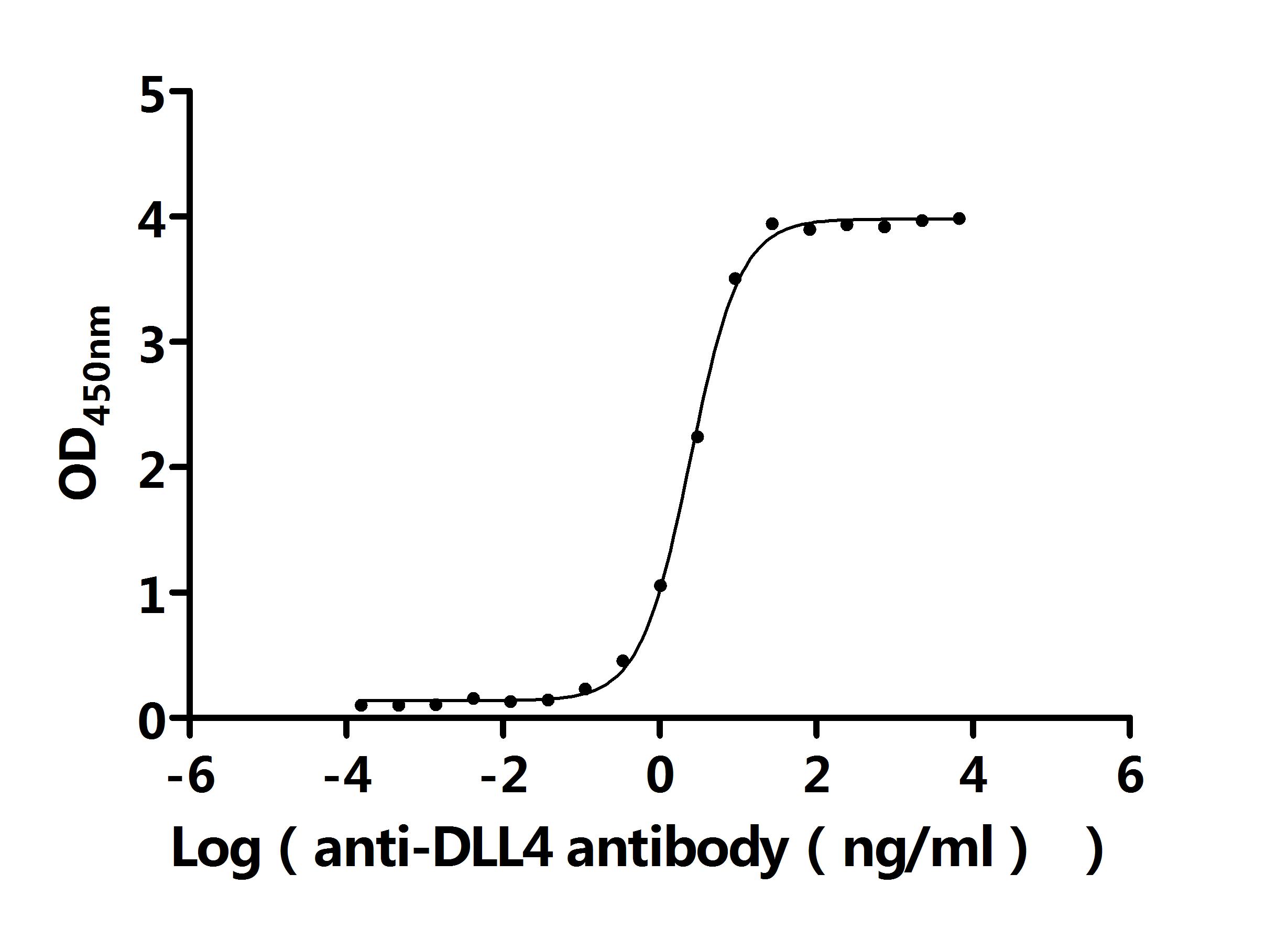
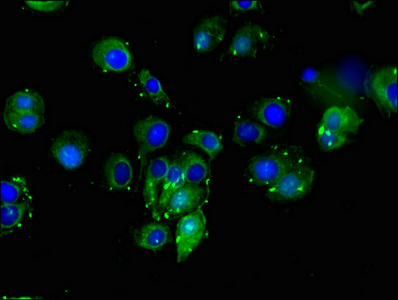
Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
Recently, Co.unlink Medicines and Compass Therapeutics announced that the phase II/III clinical trials of the VEGF/DLL4 bispecific antibody tovecimig (CTX-009/ES104) for second-line cholangiocarcinoma treatment have reached the primary endpoints. The tovecimig-based combination regimen is expected to become a new standard for second-line cholangiocarcinoma treatment globally, bringing new hope to patients.
Delta-like ligand 4 (DLL4), a key ligand in the Notch signaling pathway, plays a central role in angiogenesis and vascular remodeling. Aberrant expression of DLL4 is closely linked to abnormal angiogenesis in various pathological conditions, such as tumors, retinopathies, and cardiovascular diseases. Research indicates that DLL4 is not only a critical regulator of angiogenesis but also may contribute to disease progression by influencing endothelial cell behavior, making it a hot spot in disease therapy research in recent years.
DLL4 inhibits excessive sprouting of endothelial cells by activating Notch signaling, maintaining vascular branch balance. Under physiological conditions, DLL4 is predominantly highly expressed in tip cells of vascular endothelial cells. Through paracrine action, it binds to Notch receptors on adjacent cells, suppressing their tip cell characteristics and promoting stalk cell differentiation. This maintains vascular branch balance and prevents excessive vascular branching [2]. However, under pathological conditions, such as in the tumor microenvironment, DLL4 expression is upregulated. On one hand, by activating the Notch signaling pathway, DLL4 can inhibit angiogenesis. On the other hand, abnormal activation of this signaling may lead to aberrant vascular structures, thereby promoting tumor progression [1].
DLL4 expression is regulated by multiple transcription factors and epigenetic mechanisms. For example, in diabetic models, glucocorticoids downregulate KLF13 by inhibiting MYOD1-mediated transcriptional activation, indirectly promoting DLL4 expression. Additionally, the ubiquitination pathway, such as FBXW7-mediated degradation, regulates DLL4 protein levels, affecting its intracellular stability and function [9].
Uremic toxin indoxyl sulfate activates the OATP2B1-DLL4 axis in renal tubular epithelial cells, inducing macrophage pro-inflammatory polarization and aggravating renal interstitial fibrosis and dysfunction.
In diabetic and glucocorticoid-induced muscle atrophy models, KLF13 deficiency causes excessive DLL4-Notch2 signaling, inhibiting myocyte differentiation and promoting muscle protein degradation. Activating KLF13 or inhibiting DLL4 improves muscle quality and function [9].
DLL4 gene mutations cause hereditary capillary malformations, characterized by abnormal skin and mucosal vascular proliferation, highlighting DLL4's role in maintaining vascular homeostasis [6].
DLL4 is a key Notch signaling ligand regulating angiogenesis and vascular maturation. Anti-DLL4 antibodies can suppress tumor angiogenesis but may induce toxicities like hepatic vascular sarcomas with long-term use. Combining anti-DLL4 antibodies with anti-VEGF drugs enhances antitumor effects in mouse models and reduces abnormal vascular perfusion and drug resistance risks. Two DLL4 × VEGF bispecific antibody pipelines are currently under research, with the highest stage at phase II/III. Below is a table of ongoing pipelines:
| Drug | Mechanism of Action | Drug Type | Indications | Developers | Development Stage |
|---|---|---|---|---|---|
| Tovecimig | DLL4 inhibitor and VEGF-A inhibitor | Bispecific antibody | gallbladder cancer, intrahepatic cholangiocarcinoma, cholangiocarcinoma, colorectal cancer, metastatic colorectal cancer, rectal cancer, ovarian cancer | Compass Therapeutics LLC, et al. | Phase II/III |
| Navicixizumab | DLL4 inhibitor and VEGF inhibitor | Bispecific antibody | Advanced malignant solid tumors, colorectal cancer, ovarian cancer, gastric cancer, triple-negative breast cancer, etc. | Oncxerna Therapeutics, Inc., et al. | Phase II |
| Anti-DLL4 antibody (Regeneron) | DLL4 inhibitor | Antibody | Graft-versus-host disease | Regeneron Pharmaceuticals, Inc. | Preclinical |
CUSABIO provides high-activity DLL4 recombinant proteins, antibodies, and ELISA kits to support researchers in studying DLL4 mechanisms and exploring its clinical potential.
● DLL4 Recombinant Proteins
● DLL4 Antibodies
● DLL4 ELISA Kits
Human Delta-like protein 4(DLL4) ELISA kit
CSB-EL006949HU
References
[1] Yan M, Callahan CA, Beyer JC, et al. Chronic DLL4 blockade induces vascular neoplasms. Nature. 2010;463(7283):E6-E7. DOI: 10.1038/nature08751.
[2] Pitulescu ME, Schmidt I, Giaimo BD, et al. Dll4 and Notch signalling couples sprouting angiogenesis and artery formation. Nat Cell Biol. 2017;19(8):915-927. DOI: 10.1038/ncb3555.
[3] Folkman J. Angiogenesis: an organizing principle for drug discovery. Nat Rev Drug Discov. 2007;6(4):273-286. DOI: 10.1038/nrd2273.
[4] Bassel-Duby R, Olson EN. Notch signaling in skeletal muscle development and disease. Cold Spring Harb Perspect Biol. 2015;7(10):a019067. DOI: 10.1101/cshperspect.a019067.
[5] Yang JM, Park CS, Kim SH, et al. Dll4 Suppresses Transcytosis for Arterial Blood-Retinal Barrier Homeostasis. Circ Res. 2020;126(6):767-783. DOI: 10.1161/CIRCRESAHA.119.316476.
[6] National Cancer Institute. DLL4 wt Allele. NCI Thesaurus. Published February 8, 2020. DOI: 10.32388/QULVH4.
[7] Hellström M, Phng LK, Hofmann JJ, et al. Dll4 signaling through Notch1 regulates formation of tip cells during angiogenesis. Nature. 2007;445(7129):776-780. DOI: 10.1038/nature05571.
[8] Ren JS, Bai W, Ding JJ, et al. Hypoxia-induced AFAP1L1 regulates pathological neovascularization via the YAP-DLL4-NOTCH axis. J Transl Med. 2023;21(1):651. DOI: 10.1186/s12967-023-04503-x.
[9] Yang S, Xiong LJ, Yang GY, et al. KLF13 restrains Dll4-muscular Notch2 axis to improve muscle atrophy. J Cachexia Sarcopenia Muscle. 2024;15(5):1869-1882. DOI: 10.1002/jcsm.13538.
[10] Noguera-Troise I, Daly C, Papadopoulos NJ, et al. Blockade of Dll4 inhibits tumor growth by promoting nonproductive angiogenesis. Nature. 2006;444(7121):1032-1037. DOI: 10.1038/nature05329.
[11] Jacobsen KS, Holm T, Knoop C, et al. Notch and DLL4 expression in bevacizumab-treated colon cancer patients. Br J Cancer. 2012;107(12):1987-1993. DOI: 10.1038/bjc.2012.628.
[12] Jacobsen KS, Holm T, Knoop C, et al. Notch and DLL4 expression in bevacizumab-treated colon cancer patients [erratum]. Br J Cancer. 2013;108(3):670. DOI: 10.1038/bjc.2012.736.
[13] Li Y, Zhang Y, Wang X, et al. Palmitic acid in type 2 diabetes mellitus promotes atherosclerotic plaque vulnerability via macrophage Dll4 signaling. Atherosclerosis. 2021;327:134-144. DOI: 10.1016/j.atherosclerosis.2021.09.014.


Comments
Leave a Comment