
Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
T cell activation generally follows the classical "two-signal model": after the T cell receptor (TCR) recognizes an antigen, a second signal from co-stimulatory molecules is required to allow naive T cells to exit the quiescent state and mount an effective immune response. In this process, CD28 is one of the most critical co-stimulatory receptors [1]. As a member of the immunoglobulin superfamily, CD28 binds to B7 family molecules on the surface of antigen-presenting cells, providing the necessary signal for effective T cell activation. It not only amplifies TCR signaling, but also independently shapes downstream biological outcomes, including cytoskeleton remodeling, production of cytokines such as IL-2, enhanced proliferation, improved survival, and initiation of differentiation programs [1]. More importantly, CD28-mediated signaling can induce epigenetic, transcriptional, and post-translational changes that cannot be replicated by TCR stimulation alone [1].
1. The biological significance of CD28
2. Molecular structure, ligand recognition, and signal transduction mechanism of CD28
3. The role of CD28 in immunoregulation and disease development
The significance of CD28 is not limited to naive and effector T cells. It is also an important molecule for the maintenance and functional stability of regulatory T cells (Tregs), placing it at the intersection of immune activation and immune tolerance [1]. Precisely because of this, CD28 has obvious therapeutic development value but also brings higher regulatory complexity. Previous clinical interventions targeting CD28 family interactions have both successful experiences and obvious setbacks, indicating that although CD28 is a powerful immunoregulatory target, its role cannot be simply summarized as "enhancement" or "inhibition" [1]. Functional differences among different T cell subsets, network differences between mice and humans, and receptor-ligand crosstalk within the CD28 family all suggest that therapeutic development must emphasize temporal, cell type-specific, and context-dependent regulation [1]. Therefore, the core value of CD28 research lies not only in elucidating the basic principles of T cell co-stimulation, but also in providing a theoretical basis for precise immunoregulation and safe translation.
CD28 is a key co-stimulatory molecule on the surface of T cells, and its structure can be summarized into three functional regions: an extracellular domain responsible for ligand recognition, a single transmembrane helix, and a short intracellular tail that converts extracellular binding events into intracellular signals. The classic ligands of CD28 are the B7 family molecules CD80 and CD86. This receptor-ligand interaction constitutes the "second signal" necessary for T cell activation and plays an irreplaceable role in physiological processes such as iNKT cell development [2,3]. In therapeutic research, soluble CD80 fusion proteins that mimic natural ligands have shown potential to restore or enhance CD28 co-stimulation in the tumor microenvironment and may balance immune activation and safety window [2].
In recent years, research on the conformational dynamics of CD28 has further expanded the understanding of its regulatory mechanisms. Relevant studies suggest that acidic phospholipids in the inner leaflet of the membrane and intracellular calcium ions act as opposing charged factors to jointly regulate the conformational state of CD28 and its transmembrane signal transmission ability[4]. This view suggests that CD28 is not a simple "on/off" receptor, but more like a molecular switch finely regulated by local membrane environment and ionic conditions, which helps explain the co-stimulation threshold and its spatiotemporal specificity [4]. However, there is still a lack of sufficient high-resolution structural and dynamic experimental evidence for how this conformational change is transmitted to downstream signaling complexes through the transmembrane region and short intracellular tail [4].
In addition, the fine specificity of the CD28 ligand binding site has also attracted attention. Molecular dynamics simulations and free energy calculations suggest that some non-canonical ligands such as β-1,3-oligosaccharides may interact with CD28 with low affinity near the B7 binding site [5]. This suggests that the surface of CD28 may have a more complex interaction interface than traditional protein-protein recognition. However, such results are currently mainly limited to computational speculation, and their in vivo physiological significance still needs to be verified by more biochemical and functional experiments [5]. Overall, existing evidence indicates that CD28 ligand recognition has a clear structural basis and is dynamically regulated by membrane environment and ionic conditions, which provides an important basis for understanding its signal initiation mechanism and druggability for drug development [2-5].
After antigen stimulation, TCR proximal signaling first manifests as tyrosine phosphorylation of CD3 and TCRζ chains, and recruitment and activation of Lck and ZAP70. Live-cell TIRF imaging studies show that CD28 co-stimulation can significantly accelerate the recruitment of Lck to TCR microclusters and change the spatial distribution of active and inactive Lck, thereby promoting faster and more sufficient binding and activation of ZAP70, ultimately lowering the TCR activation threshold and enhancing the proximal signaling amplification effect of CD3ζ [6]. This result indicates that CD28 does not only enhance signaling downstream, but reshapes the signaling microenvironment very early to change T cell activation efficiency [6].
On this basis, CD28 further transmits signals to multiple classical CD28 signaling pathways. A computational and experimental CAR-T model suggests that CD28 can accelerate ERK activation by enhancing CD3ζ phosphorylation kinetics, indicating that its impact on the MAPK pathway may mainly lie in amplifying proximal tyrosine kinase events rather than directly initiating downstream MAPK modules [7]. Phosphorylation profiling further shows that CD28 co-stimulation can induce a specific set of tyrosine kinase and receptor tyrosine kinase phosphorylation patterns, indicating that different co-stimulation pathways have molecular differences in the selection of signaling nodes [8]. Meanwhile, CD28 can also coordinate PLCγ-1/IP3R-related calcium signals, thereby activating transcription factors such as NFAT, AP-1, and NF-κB, and promoting the expression of IL-2 and other cytokines [9].CD28 signaling is also constrained by a variety of negative regulatory pathways. TIM-3 can interact with Lck, ITK, and the PI3K adaptor p85, thereby inhibiting CD3/CD28-induced NF-κB and NFAT activities and reducing IL-2 secretion[10]. B7-H7 (HHLA2) exhibits an inhibitory effect when co-stimulated with TCR and CD28, and its transcriptional response is significantly weaker than classical B7-1 co-stimulation, suggesting that it can play a "braking" role in physiological APC-T cell interactions [11]. More notably, Shp2 recruited by PD-1 has a stronger preference for dephosphorylation of CD28 than TCR components, indicating that immune checkpoints can weaken T cell function by directly inhibiting CD28 co-stimulation [12]. These findings have changed the understanding of co-inhibitory signaling and highlight the central role of CD28 in maintaining effector T cell function.
In addition to the classical kinase-transcription axis, CD28 also affects T cell fate through post-transcriptional regulation and metabolic reprogramming. In CD8+ T cells, CD28 can upregulate ARS2, promote the recruitment of splicing factors, and alter activation-related alternative splicing events. The splicing switch from PKM1 to PKM2 has been shown to be closely related to glucose metabolic flexibility, IFN-γ production, and antitumor function, and this process is independent of the PI3K pathway driven by CD28 [13]. Meanwhile, under certain conditions, CD28 can also inhibit cell adhesion by mediating Rap1 inactivation through CAPRI, indicating that its signal output is not unidirectional enhancement, but has obvious context dependence and functional differentiation [14]. Therefore, CD28 should be understood as a complex signaling node integrating proximal kinases, transcription factors, alternative splicing, and metabolic programs, rather than a single-path amplifier [6-14].
During the establishment of immune tolerance, B7-CD28 co-stimulation is involved in both clonal deletion in the thymus and supports Treg development, but the degree of dependence on CD28 signaling and the demand for different antigen-presenting cells are not the same [15]. Mouse studies show that both clonal deletion and Treg induction of endogenous tissue-restricted antigen-specific thymocytes depend on B7-CD28 interaction, but there are differences in signal intensity, time window, and APC source requirements [15]. More importantly, when abnormal B7-CD28 signaling impairs thymic clonal deletion, mature autoreactive T cells can accumulate in the periphery and acquire the ability to mediate destructive autoimmunity [15]. This indicates that CD28 is not only involved in effector activation, but also deeply involved in the establishment of central tolerance.
Clinical intervention evidence also suggests that CD28 signaling imbalance is closely related to autoimmunity. Therapy targeting the CTLA-4/B7 axis can affect T cell responses by changing the opportunity for CD28 to obtain co-stimulation, which has therapeutic value in some contexts but may also disrupt Treg homeostasis and tolerance maintenance. Thus, the CD28 pathway is not suitable for simple global enhancement or global inhibition, but requires precise regulation according to disease background, T cell subset composition, and pathological stage [15-17].
In tumor immunity, CD28 first acts as a key co-stimulatory molecule required for anti-tumor responses. Conditional knockout and chronic infection/tumor models both show that CD28/B7 signaling is important for maintaining CD8+ T cell function and improving therapeutic responses such as anti-PD-1 [18]. This indicates that even in the context of immune checkpoint blockade, effector T cells still need CD28 to provide co-stimulatory support to maintain strong killing and proliferative capacity [18].
However, the role of CD28 is not entirely unidirectional pro-effector. With the deepening of research, it has been found that CD28 function is affected by the complex influence of tumor microenvironment, cell type, and spatial location. For example, some studies suggest that the tumor-associated environment can reshape CD28 output by changing the balance between co-stimulation and co-inhibition signals, and even limit therapeutic benefits in some cases. In addition, in vivo CRISPR screening also suggests that in specific tumor models, cancer cells themselves may have more complex non-classical connections with CD28-related pathways, which means that the function of CD28 in tumors should not only be understood from the perspective of T cells [19-21]. Therefore, the value of CD28 in tumor immunity is not only reflected in promoting anti-tumor T cell responses, but also in being an important intersection connecting co-stimulation, checkpoints, and microenvironmental suppression.
With age, the peripheral immune system undergoes significant remodeling, among which the decrease in CD28 expression is one of the most representative characteristics of T cell aging. Cross-sectional analysis of a large population shows that increasing age is accompanied by a decrease in naive T cells and an increase in the proportion of terminally differentiated or senescence-associated T cells, while the decrease in CD28 expression suggests that the basis for T cells to obtain effective co-stimulation and proliferation is weakened [22]. Functionally, this change may lead to decreased primary immune response ability, weakened immune surveillance, and persistent chronic low-grade inflammation.
Although most of these studies are cross-sectional design, it is difficult to directly infer causal changes in individual life cycles, existing evidence suggests that CD28 loss is significantly associated with immunosenescence, "inflammaging", and increased risk of certain chronic diseases [22,23]. Therefore, CD28 is not only a T cell co-stimulatory molecule, but also an important indicator for observing immunosenescence and immune function decline.
Currently, CD28 targeted drug development is active, covering various types such as CAR-T, bispecific/multispecific antibodies, mainly used in autoimmune diseases and various cancers. Some products, such as those for lupus nephritis and lymphoma, have entered phase 2/3 and phase 2 clinical trials, and more than 30 other products are in phase 1/2 or phase 1, showing broad therapeutic prospects. Part of the pipeline is listed in the table below:
| Medications | Target | Types of medications | Indications under investigation | Research institutions | Highest R&D stage |
|---|---|---|---|---|---|
| Mivocabtagene Autoleucel(Kyverna) | CD19 x CD28 | Autologous CAR-T | Lupus nephritis, Rheumatoid arthritis, Systemic scleroderma, Stiff-person syndrome, Myasthenia gravis, etc. | Kyverna Therapeutics, Inc., Hannover Medical School | Phase 2/3 |
| CD19CAR-CD28-CD3zeta-EGFRt-expressing Tn/mem-enriched T-lymphocytes(City of Hope National Medical Center) | CD19 x CD28 x EGFR | Autologous CAR-T | Relapsed mantle cell lymphoma, Refractory mantle cell lymphoma, Primary central nervous system lymphoma, etc. | City of Hope National Medical Center | Phase 2 |
| Pegrizeprument | CD28 | Fab fragment antibody | Renal transplant rejection, Organ transplant rejection, Rheumatoid arthritis | Veloxis Pharmaceuticals A/S, Biomedical Primate Research Centre, OSE Immunotherapeutics SA, Janssen Global Services LLC | Phase 2 |
| Acazicolcept | CD28 x ICOS | Bispecific antibody | Lupus nephritis, Systemic lupus erythematosus | Alpine Immune Sciences, Inc. | Phase 2 |
| Dalmitamig | CD28 x EGFR | Bispecific antibody | Metastatic non-small cell lung cancer, Stage IIIB non-small cell lung cancer, Locally advanced rectal cancer, etc. | Regeneron Pharmaceuticals, Inc. | Phase 2 |
| REGN-5668 | CD28 x MUC16 | Bispecific antibody | Platinum-resistant primary peritoneal cancer, Recurrent ovarian cancer, Endometrial cancer, etc. | Regeneron Pharmaceuticals, Inc. | Phase 1/2 |
| SIM-323 | CD28 x CTLA4 x IL-2Rβγ | Fc fusion protein | Advanced malignant solid tumors, Bladder cancer, Colorectal cancer, Esophageal squamous cell carcinoma, etc. | GI Innovation, Inc. | Phase 1/2 |
| REGN-17372 | CD28 x GPRC5D | Bispecific antibody | - | Regeneron Pharmaceuticals, Inc. | Phase 1/2 |
| ZGGS34 | CD28 x CD3 x MUC17 | Trispecific T cell engager | Advanced malignant solid tumors | Suzhou Zelgen Biopharmaceuticals Co., Ltd. | Phase 1/2 |
| MDX-2003 | CD19 x CD20 x CD28 x CD3 | Tetraspecific antibody | Follicular lymphoma, High-grade B-cell lymphoma, Lymphoplasmacytic lymphoma, etc. | ModeX Therapeutics, Inc. | Phase 1/2 |
| Nezastomig | CD28 x PSMA | Bispecific antibody | Advanced prostate cancer, Metastatic castration-resistant prostate cancer, Renal cell carcinoma | Regeneron Pharmaceuticals, Inc. | Phase 1/2 |
| MDX-2001 | CD28 x CD3 x Trop-2 x c-Met | Tetraspecific T cell engager | Advanced malignant solid tumors | ModeX Therapeutics, Inc. | Phase 1/2 |
| REGN-7945 | CD28 x CD38 | Bispecific antibody | Relapsed multiple myeloma | Regeneron Pharmaceuticals, Inc. | Phase 1/2 |
| SAR-446422 | CD28 x OX40 | Bispecific antibody | Inflammation | Sanofi | Phase 1 |
| CD19/CD28 CAR-T (Baylor College of Medicine) | CD19 x CD28 | Autologous CAR-T | CD19-expressing malignancies, Chronic lymphocytic leukemia, Precursor B-cell lymphoblastic leukemia lymphoma | Baylor College of Medicine | Phase 1 |
| JNJ-1493 | CD20 x CD28 | Bispecific antibody | Non-Hodgkin lymphoma, Diffuse large B-cell lymphoma, High-grade B-cell lymphoma, etc. | Janssen Research & Development LLC, Janssen-Cilag International NV, Xencor, Inc., Johnson & Johnson | Phase 1 |
| RNDO-564 | CD28 x nectin-4 | Bispecific antibody | Bladder cancer, Esophageal squamous cell carcinoma, Adenocarcinoma of the gastroesophageal junction, Non-small cell lung cancer, etc. | Rondo Therapeutics, Inc. | Phase 1 |
| Allogeneic anti-CD28|CD5 CAR-T (Baylor College of Medicine) | CD28 x CD5 | Off-the-shelf CAR-T | Adult T-cell leukemia/lymphoma, Anaplastic large cell lymphoma, Enteropathy-associated T-cell lymphoma, etc. | Baylor College of Medicine | Phase 1 |
| SSS59 | CD28 x CD3 x MUC17 | Trispecific T cell engager | Advanced cancer | Shenyang Sansheng Pharmaceutical Co., Ltd. | Phase 1 |
| HLX-3901 | CD28 x CD3 x DLL3 | Tetraspecific T cell engager | Neuroendocrine carcinoma, Small cell carcinoma, Small cell lung cancer, Advanced malignant solid tumors, etc. | Shanghai Fosun RuiLin Biotechnology Co., Ltd., Shanghai Fosun Hanlin Biotechnology Co., Ltd., Shanghai Fosun Hanlin Biomedical Co., Ltd. | Phase 1 |
| BI-1821736 | CD28 | Oncolytic virus | Advanced cancer, Advanced malignant solid tumors | Boehringer Ingelheim España SA | Phase 1 |
| JNJ-87189401 | CD28 x PSMA | Bispecific antibody | Castration-resistant prostate cancer, Recurrent prostate cancer, Prostate adenocarcinoma, etc. | Janssen-Cilag International NV, Xencor, Inc., Janssen Research & Development LLC | Phase 1 |
| REGN-5837 | CD22 x CD28 | Bispecific antibody | Diffuse large B-cell lymphoma, Relapsed non-Hodgkin lymphoma, Aggressive B-cell non-Hodgkin lymphoma | Regeneron Pharmaceuticals, Inc. | Phase 1 |
| SG-1827 | CD28 x CTLA4 | Fc fusion protein | Advanced malignant solid tumors | Hangzhou Sagen Biotechnology Co., Ltd. | Phase 1 |
| RG-6333 | CD19 x CD28 | Bispecific antibody | Refractory B-cell lymphoma | F. Hoffmann-La Roche Ltd. | Phase 1 |
| iC9-GD2-CD28-OX40(Baylor College of Medicine) | CD28 x GD2 x OX40 | CAR-T | Refractory neuroblastoma | Baylor College of Medicine | Phase 1 |
| XmAb-808 | CD276 x CD28 | Bispecific antibody | Advanced malignant solid tumors, Colorectal cancer, Melanoma, Metastatic castration-resistant prostate cancer, etc. | Xencor, Inc. | Phase 1 |
| Anti-CD28|CD7 CAR-T (Baylor College of Medicine) | CD28 x CD7 | Autologous CAR-T | Adult T-cell leukemia/lymphoma, Anaplastic large cell lymphoma, Enteropathy-associated T-cell lymphoma, etc. | Baylor College of Medicine | Phase 1 |
| CHM-1101 | CD19 x CD28 x CD3 | Autologous CAR-T | Glioblastoma multiforme, MMP2-positive glioblastoma, Recurrent glioblastoma, etc. | Chimeric Therapeutics Ltd. (Australia), City of Hope National Medical Center, Chimeric Therapeutics Ltd. | Phase 1 |
| CC-312 | CD19 x CD28 x CD3 | Trispecific T cell engager | Systemic lupus erythematosus, Aggressive B-cell non-Hodgkin lymphoma, Refractory B-cell lymphoma, etc. | Huihe Biotechnology (Shanghai) Co., Ltd. | Phase 1 |
(Data cutoff: March 15, 2026, sourced from synapse)
CD28 is a core molecule in the T cell co-stimulation network, and plays an irreplaceable role in T cell activation, proliferation, metabolic reprogramming, effector maintenance, and immune tolerance. It has developed from a classic T cell co-stimulatory molecule to an important target with both in-depth mechanism research and clinical translation potential. CUSABIO provides CD28 recombinant proteins, antibodies, and ELISA kits to assist you in related mechanism research and targeted drug development.
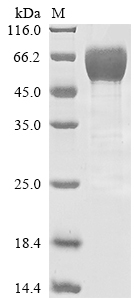
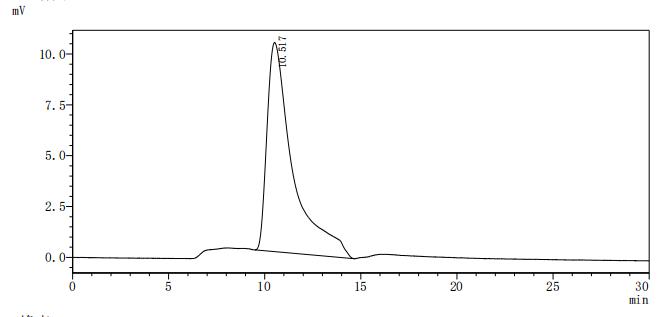
● CD28 Recombinant Proteins
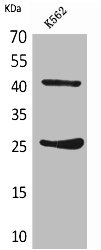
● CD28 Antibodies
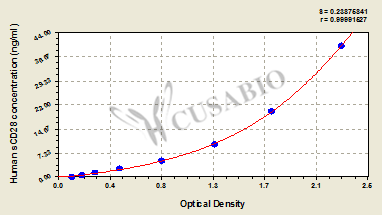
● CD28 ELISA Kits
References
[1] Jonathan H Esensten, Ynes A Helou, Gaurav Chopra, Arthur Weiss, Jeffrey A Bluestone.(2016). CD28 Costimulation: From Mechanism to Therapy.
[2] Songna Wang, Pinliang Hu, Xuyao Zhang, Jiajun Fan, Jing Zou, Weidong Hong, Xuan Huang, Danjie Pan, Huaning Chen, Dianwen Ju, Yi Zhun Zhu, Li Ye.(2025). Recombinant CD80 fusion protein combined with discoidin domain receptor 1 inhibitor for cancer treatment.
[3] Masashi Watanabe, Sofia Celli, Farrah A Alkhaleel, Richard J Hodes.(2022). Antigen-presenting T cells provide critical B7 co-stimulation for thymic iNKT cell development via CD28-dependent trogocytosis.
[4] Wei Yang, Weiling Pan, Shuokai, Nicola Trendel, Shutan Jiang, Feng Xiao, Manman Xue, Wei Wu, Zeli Peng, Xiaoxi Li, Hongbin Ji, Xiaolong Liu, Hai Jiang, Haopeng Wang, Hongbin Shen, Omer Dushek, Hua Li, Chenqi Xu.(2017). Dynamic regulation of CD28 conformation and signaling by charged lipids and ions.
[5] Jeffrey Comer, Molly Bassette, Riley Burghart, Mayme Loyd, Susumu Ishiguro, Ettayapuram Ramaprasad Azhagiya Singam, Ariela Vergara-Jaque, Ayaka Nakashima, Kengo Suzuki, Brian V Geisbrecht, Masaaki Tamura.(2021). Beta-1,3 Oligoglucans Specifically Bind to Immune Receptor CD28 and May Enhance T Cell Activation.
[6] Kumarkrishna Raychaudhuri, Rohita Rangu, Alison Ma, Neriah Alvinez, Andy D Tran, Sandeep Pallikkuth, Katherine M McIntire, Joseph A Garvey, Jason Yi, Lawrence E Samelson.(2024). CD28 shapes T cell receptor signaling by regulating Lck dynamics and ZAP70 activation.
[7] Jennifer A. Rohrs, Elizabeth L. Siegler, Pin Wang, Stacey D. Finley.(2020). ERK Activation in CAR T Cells Is Amplified by CD28-Mediated Increase in CD3ζ Phosphorylation.
[8] Anuja Bhatta, Marcia A Chan, Stephen H Benedict.(2021). Transcription factor activation and protein phosphorylation patterns are distinct for CD28 and ICAM-1 co-stimulatory molecules.
[9] Xuelian Xiang, Nan Cao, Feiyue Chen, Long Qian, Yifei Wang, Yunmao Huang, Yunbo Tian, Danning Xu, Wanyan Li.(2020). Polysaccharide of Atractylodes macrocephala Koidz (PAMK) Alleviates Cyclophosphamide-induced Immunosuppression in Mice by Upregulating CD28/IP3R/PLCγ-1/AP-1/NFAT Signal Pathway.
[10] Brian Tomkowicz, Eileen S. Walsh, Adam Cotty, Raluca Verona, Nina Chi-Sabins, Fred M. Kaplan, Sandy Santulli-Marotto, Chen-Ni Chin, Jill M. Mooney, Russell B. Lingham, Michael Naso, Timothy J. McCabe.(2015). TIM-3 Suppresses Anti-CD3/CD28-Induced TCR Activation and IL-2 Expression through the NFAT Signaling Pathway.
[11] Sadiye Amcaoglu Rieder, Jingya Wang, Natalie White, Ariful Qadri, Catherine Menard, Geoffrey Stephens, Jodi L Karnell, Christopher E Rudd, Roland Kolbeck.(2021). B7-H7 (HHLA2) inhibits T-cell activation and proliferation in the presence of TCR and CD28 signaling.
[12] Enfu Hui, Jeanne Cheung, Jing Zhu, Xiaolei Su, Marcus J Taylor, Heidi A Wallweber, Dibyendu K Sasmal, Jun Huang, Jeong M Kim, Ira Mellman, Ronald D Vale.(2017). T cell costimulatory receptor CD28 is a primary target for PD-1-mediated inhibition.
[13] G Aaron Holling, Colin A Chavel, Anand P Sharda, Mackenzie M Lieberman, Caitlin M James, Shivana M Lightman, Jason H Tong, Guanxi Qiao, Tiffany R Emmons, Thejaswini Giridharan, Shengqi Hou, Andrew M Intlekofer, Richard M Higashi, Teresa W M Fan, Andrew N Lane, Kevin H Eng, Brahm H Segal, Elizabeth A Repasky, Kelvin P Lee, Scott H Olejniczak.(2024). CD8+ T cell metabolic flexibility elicited by CD28-ARS2 axis-driven alternative splicing of PKM supports antitumor immunity.
[14] Marianne Strazza, Inbar Azoulay-Alfaguter, Bryan Dun, Jairo Baquero-Buitrago, Adam Mor.(2015). CD28 inhibits T cell adhesion by recruiting CAPRI to the plasma membrane.
[15] Masashi Watanabe, Ying Lü, Michael Breen, Richard J. Hodes.(2020). B7-CD28 co-stimulation modulates central tolerance via thymic clonal deletion and Treg generation through distinct mechanisms.
[16] Yinghao Zhao, Wei Yang, Yuanyuan Huang, Ranji Cui, Xiangyan Li, Bingjin Li.(2018). Evolving Roles for Targeting CTLA-4 in Cancer Immunotherapy.
[17] Shunsuke Fukuyo, Shingo Nakayamada, Shigeru Iwata, Satoshi Kubo, Kazuyoshi Saito, Yoshiya Tanaka.(2017). Abatacept therapy reduces CD28+CXCR5+ follicular helper-like T cells in patients with rheumatoid arthritis.
[18] Alice O Kamphorst, Andreas Wieland, Tahseen Nasti, Shu Yang, Ruan Zhang, Daniel L Barber, Bogumila T Konieczny, Candace Z Daugherty, Lydia Koenig, Ke Yu, Gabriel L Sica, Arlene H Sharpe, Gordon J Freeman, Bruce R Blazar, Laurence A Turka, Taofeek K Owonikoko, Rathi N Pillai, Suresh S Ramalingam, Koichi Araki, Rafi Ahmed.(2017). Rescue of exhausted CD8 T cells by PD-1-targeted therapies is CD28-dependent.
[19] Wei Huff, Marpe Bam, Jack Shireman, Jae Hyun Kwon, Leo Song, Sharlé Newman, Aaron Cohen‐Gadol, Scott Shapiro, Tamara Jones, Kelsey Fulton, Sheng Liu, Hiromi Tanaka, Yunlong Liu, Jun Wan, Mahua Dey.(2021). Aging- and Tumor-Mediated Increase in CD8+CD28− T Cells Might Impose a Strong Barrier to Success of Immunotherapy in Glioblastoma.
[20] Zhen Yang, Xinpeng Liu, Jun Zhu, Yangyang Chai, Boyi Cong, Bo Li, Wanfeng Gao, Ye Hu, Mingyue Wen, Yanfang Liu, Li Fu, Xuetao Cao.(2025). Inhibiting intracellular CD28 in cancer cells enhances antitumor immunity and overcomes anti-PD-1 resistance via targeting PD-L1.
[21] Ryan C Augustin, Riyue Bao, Jason J Luke.(2023). Targeting Cbl-b in cancer immunotherapy.
[22] Xuexiao Jin, Qin Xu, Chengfei Pu, Kaixiang Zhu, Cheng Lu, Yu Jiang, Lei Xiao, Yongmei Han, Linrong Lu.(2021). Therapeutic efficacy of anti-CD19 CAR-T cells in a mouse model of systemic lupus erythematosus.
[23] Ling Qin, Jing Xie, Zhifeng Qiu, Wei Cao, Yang Jiao, Jean‐Pierre Routy, Taisheng Li.(2016). Aging of immune system: immune signature from peripheral blood lymphocyte subsets in 1068 healthy adults.


Comments
Leave a Comment