Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
The Notch signaling pathway plays a pivotal role in biological systems, regulating not only embryonic development and adult tissue homeostasis but also exerting dual functions in the initiation and progression of multiple cancers - acting as either a tumor suppressor or a driver of cancer cell proliferation, angiogenesis, and immune evasion. This dedicated resource systematically summarizes core knowledge on Notch1 - 4 receptors and DLL/JAG ligands, enabling you to rapidly identify high-quality recombinant proteins, antibodies, and ELISA kits to advance your research in oncology and other Notch-related disease areas.
The Notch signaling pathway is an evolutionarily conserved cell-to-cell communication system, first identified through the "notched" wing phenotype in Drosophila. In mammals, it consists of four transmembrane receptors (Notch1–4) and five canonical ligands (DLL1, DLL3, DLL4, JAG1, JAG2), activated exclusively upon direct contact between adjacent cells to precisely regulate cell fate determination, differentiation, proliferation, and tissue homeostasis.
All Notch receptors are type I transmembrane proteins. They are cleaved by furin in the Golgi apparatus and trafficked to the cell membrane as heterodimers. Ligands are also transmembrane proteins expressed on the surface of neighboring cells. Upon ligand-receptor binding, two sequential proteolytic cleavages occur: an S2 cleavage mediated by ADAM metalloproteases, followed by an S3 cleavage catalyzed by γ-secretase. This releases the Notch intracellular domain (NICD), which translocates to the nucleus, forms a transcriptional complex with RBP-J and other co-factors, and activates target genes such as Hes and Hey, thereby transmitting the signal.
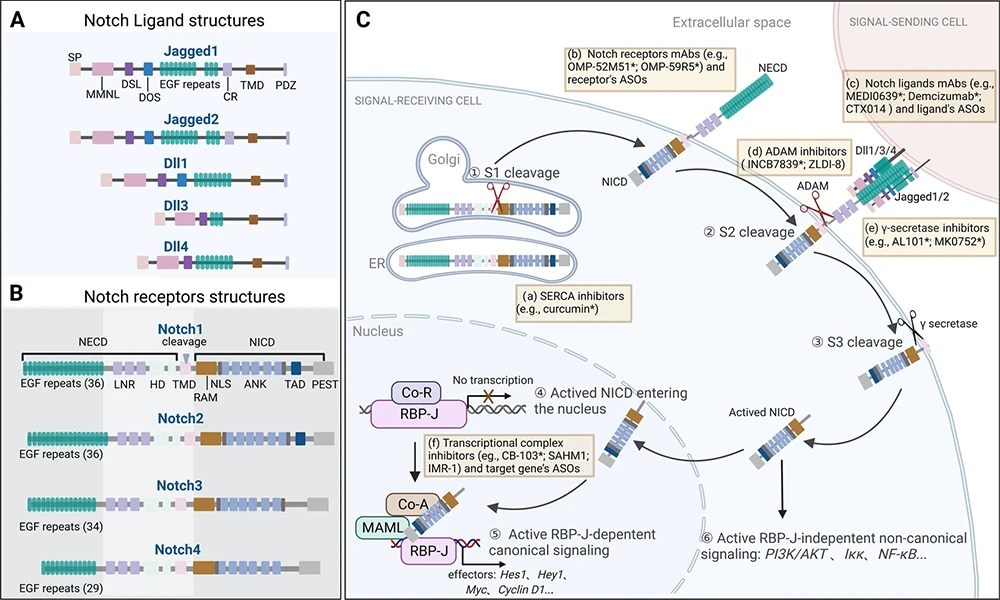
Figure: NOTCH Signaling Pathway [1]
(A. Ligand structural features; B. Receptor structural features; C. Signal reception and transduction process.)
Although the core mechanism is highly conserved, distinct Notch receptors and ligands exhibit significant differences in tissue distribution, binding preferences, and biological functions—for example, DLL4–Notch1 dominates angiogenesis, JAG1–Notch2 participates in kidney development, and DLL3 is aberrantly overexpressed in small cell lung cancer (SCLC). These specificities underscore the importance of precise target selection and reagent matching in research.
The functional diversity of the Notch signaling pathway arises from the differential expression of its receptors and ligands across tissues and developmental stages, leading to context-dependent signaling outputs. The tables below summarize the expression profiles, core biological functions, and disease associations of the four Notch receptors and five canonical ligands for quick reference.
| Target | Primary Expression Sites | Core Biological Functions | Disease Associations |
|---|---|---|---|
| Notch1 | Thymic T cells, vascular endothelium, neural progenitors, mammary epithelium | T-cell development, angiogenesis, stem cell maintenance | T-cell acute lymphoblastic leukemia (T-ALL) [2,3], breast cancer [4], melanoma [5], hepatocellular carcinoma [6], colorectal cancer [7], Alzheimer’s disease [20] |
| Notch2 | Renal tubular epithelium, intrahepatic bile duct cells, B cells, bone marrow stroma | Kidney/bile duct development, B-cell differentiation, liver regeneration | Chronic lymphocytic leukemia (B-CLL) [8], small cell lung cancer [9], pancreatic cancer [10], breast cancer [11], Hajdu-Cheney syndrome [18], Alagille syndrome [19] |
| Notch3 | Vascular smooth muscle cells, cerebral arterioles, pericytes | Vascular smooth muscle homeostasis, neurovascular function | Cerebral autosomal dominant arteriopathy with subcortical infarcts and leukoencephalopathy (CADASIL) [12] |
| Notch4 | Vascular endothelium, mammary epithelium | Mammary epithelial development, endothelial barrier function | Systemic sclerosis [13], melanoma [14], hepatocellular carcinoma [15], asthma [16], bipolar disorder [17] |
| Target | Primary Expression Sites | Core Biological Functions | Disease Associations |
|---|---|---|---|
| DLL1 | Neural stem cells, somites, hematopoietic niche, intestinal crypts | Neurogenesis, somitogenesis, hematopoietic stem/progenitor cell regulation | Breast cancer [21], neuroblastoma [22], multiple myeloma [22] |
| DLL3 | Neuroendocrine tissues, fetal brain, small cell lung cancer (SCLC) | Inhibits canonical Notch signaling; regulates neuroendocrine fate | Small cell lung cancer (SCLC) [24–26], neuroendocrine carcinomas [27] |
| DLL4 | Vascular "tip cells", tumor-associated endothelium, thymic epithelium | Regulates angiogenic sprouting; restricts excessive tip cell formation | Colorectal cancer [28], breast cancer [29], lung cancer [30], renal cancer [31], gastric cancer [32] |
| JAG1 | Biliary/renal epithelium, endocardium, hepatic stellate cells, CAFs | Organogenesis (liver/kidney/heart), immune modulation, fibrosis | Alagille syndrome [33], renal fibrosis [34], triple-negative breast cancer [35] |
| JAG2 | Osteoblasts, placental trophoblasts, T cells, cardiac valves | Bone development, T-cell tolerance | Colorectal cancer [36], multiple myeloma [37], lung cancer [38] |
DLL3 is overexpressed in more than 80% of small cell lung cancer (SCLC) cases and is virtually absent in normal adult tissues, making it an ideal tumor-specific antigen.
Read more: DLL3: A New Molecular Target in Cancer
View all DLL3-related productsDLL4 acts as a negative regulator during angiogenesis—activating Notch1 on endothelial cells to suppress excessive sprouting and maintain vascular network integrity. However, in multiple solid tumors (e.g., breast, colorectal, and liver cancers), DLL4 is aberrantly overexpressed, leading to disorganized, poorly perfused tumor vasculature that promotes hypoxia, invasion, and immune evasion.
Read more: DLL4: A Key Target for Angiogenesis Regulation
View all DLL4-related productsThe Notch signaling pathway plays a central regulatory role in key biological processes including development, tissue homeostasis, and tumorigenesis. In-depth investigation of this pathway relies on highly specific and reliable experimental reagents.
CUSABIO offers a comprehensive portfolio of antibodies, recombinant proteins, and ELISA kits covering the entire Notch receptor and ligand family, empowering your research with precision and efficiency.
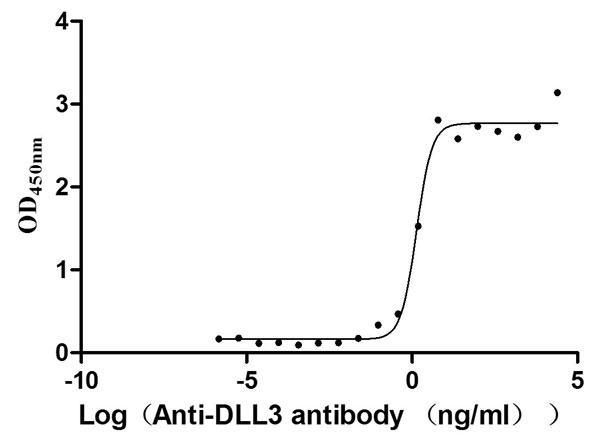
Code: CSB-RA882142MA2HU
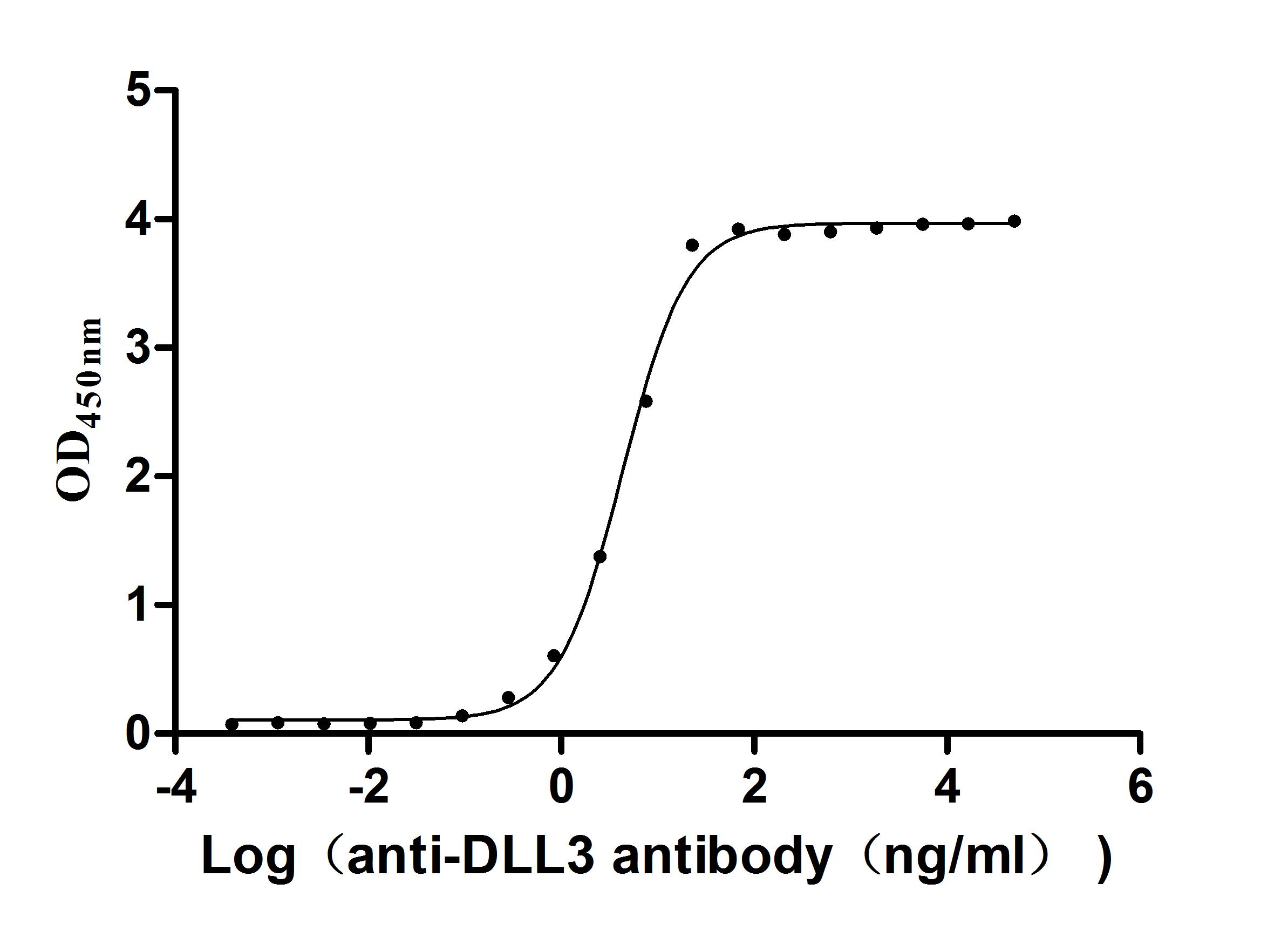
The Binding Activity of Human DLL3 with Anti-DLL3 recombinant
antibody
Activity: Measured by its binding ability in a functional ELISA.
Immobilized Human DLL3 (CSB-MP882142HU2d7) at 2 μg/mL can bind Anti-DLL3 recombinant
antibody. The EC50 is 3.990-4.723 ng/mL.
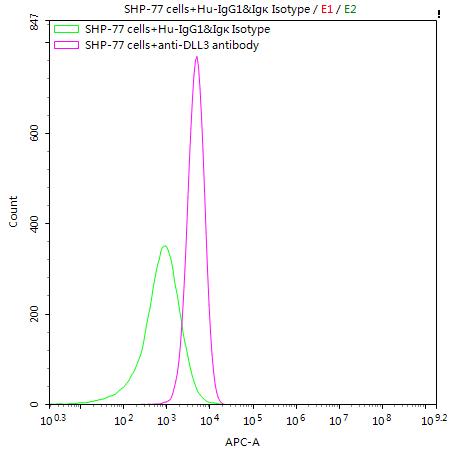
SHP-77 cells were stained with Human IgG1&Igκ Isotype Control (CSB-RA011156MA1HU)(green line) and anti-DLL3 antibody (CSB-RA882142MA2HU) (2µg/1*106cells) (red line), washed and then followed by APC-conjugated anti-Human IgG Fc antibody and analyzed with flow cytometry.
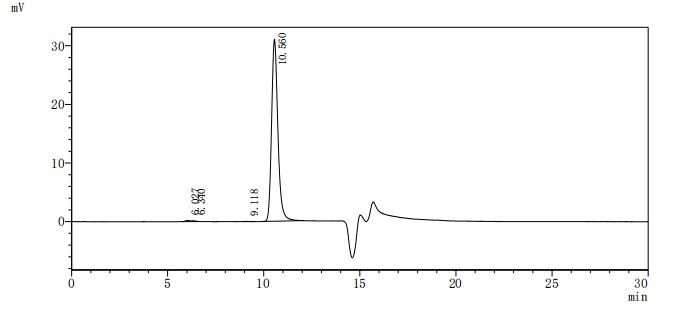
The purity of DLL3 was greater than 95% as determined by SEC-HPLC
Code: CSB-MP882142HU
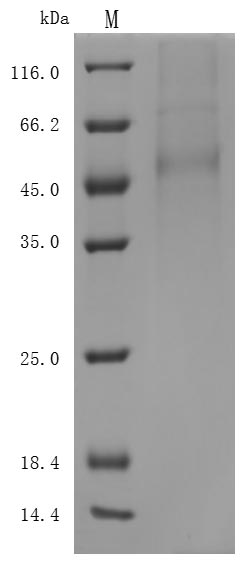
(Tris-Glycine gel) Discontinuous SDS-PAGE (reduced) with 5% enrichment gel and 15% separation gel.
Activity: Measured by its binding ability in a functional ELISA. Immobilized DLL3 at 2 μg/ml can bind Anti-DLL3 Recombinant Antibody(CSB-RA882142A1HU), the EC50 is 1.102-1.707 ng/mL.
Code: CSB-EL006948HU
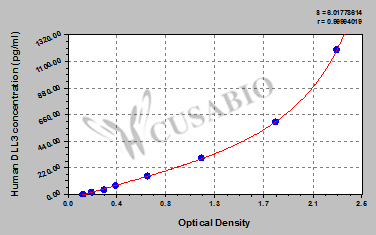
These standard curves are provided for demonstration only. A standard curve should be generated for each set of samples assayed.
| Target | Code | Product Name | Source |
|---|---|---|---|
| DLL3 | CSB-EP882142HU | Recombinant Human Delta-like protein 3 (DLL3), partial | E.coli |
| DLL3 | CSB-YP882142HU | Recombinant Human Delta-like protein 3 (DLL3), partial | Yeast |
| DLL3 | CSB-EP882142HU1 | Recombinant Human Delta-like protein 3 (DLL3), partial | E.coli |
| DLL3 | CSB-MP882142HU | Recombinant Human Delta-like protein 3 (DLL3), partial (Active) | Mammalian cell |
| DLL3 | CSB-MP3536MOV | Recombinant Macaca fascicularis Delta-like protein 3 (DLL3), partial (Active) | Mammalian cell |
| DLL3 | CSB-EP882142HU2-B | Recombinant Human Delta-like protein 3 (DLL3), partial, Biotinylated | E.coli |
| DLL3 | CSB-MP882142HU2 | Recombinant Human Delta-like protein 3 (DLL3), partial (Active) | Mammalian cell |
| DLL3 | CSB-MP882142HU2d7 | Recombinant Human Delta-like protein 3 (DLL3), partial (Active) | Mammalian cell |
| DLL3 | CSB-MP882142HU3 | Recombinant Human Delta-like protein 3 (DLL3), partial (Active) | Mammalian cell |
| DLL3 | CSB-MP882142HU3d7 | Recombinant Human Delta-like protein 3 (DLL3), partial (Active) | Mammalian cell |
| DLL4 | CSB-MP878862HU | Recombinant Human Delta-like protein 4 (DLL4), partial (Active) | Mammalian cell |
| DLL4 | CSB-MP7292MOV-C | Recombinant Macaca fascicularis Delta-like protein (DLL4), partial | Mammalian cell |
| Jag1 | CSB-EP870758MO | Recombinant Mouse Protein jagged-1 (Jag1), partial | E.coli |
| JAG1 | CSB-MP011927HUh6 | Recombinant Human Protein jagged-1 (JAG1), partial | Mammalian cell |
| NOTCH1 | CSB-EP015949HU | Recombinant Human Neurogenic locus notch homolog protein 1 (NOTCH1), partial | E.coli |
| NOTCH2NLB | CSB-BP3322HU | Recombinant Human Notch homolog 2 N-terminal-like protein B (NOTCH2NLB) | Baculovirus |
| NOTCH2NLB | CSB-EP3322HU | Recombinant Human Notch homolog 2 N-terminal-like protein B (NOTCH2NLB) | E.coli |
| Target | Code | Product Name | Tested Applications |
|---|---|---|---|
| DLL1 | CSB-PA006947GA01HU | DLL1 Antibody | ELISA, WB |
| DLL3 | CSB-RA882142MA2HU | DLL3 Recombinant Monoclonal Antibody | ELISA, FC |
| DLL3 | CSB-PA882142LA01HU | DLL3 Antibody | ELISA, IHC |
| DLL3 | CSB-PA882142LB01HU | DLL3 Antibody, HRP conjugated | ELISA |
| DLL3 | CSB-PA882142LC01HU | DLL3 Antibody, FITC conjugated | N/A |
| DLL3 | CSB-PA882142LD01HU | DLL3 Antibody, Biotin conjugated | ELISA |
| DLL4 | CSB-PA073786 | DLL4 Antibody | ELISA, IHC |
| DLL4 | CSB-PA831848 | DLL4 Antibody | ELISA, IHC |
| DLL4 | CSB-PA006949LA01HU | DLL4 Antibody | ELISA, IF |
| DLL4 | CSB-PA006949LB01HU | DLL4 Antibody, HRP conjugated | ELISA |
| DLL4 | CSB-PA006949LC01HU | DLL4 Antibody, FITC conjugated | N/A |
| DLL4 | CSB-PA006949LD01HU | DLL4 Antibody, Biotin conjugated | ELISA |
| JAG1 | CSB-RA272247A0HU | JAG1 Recombinant Monoclonal Antibody | ELISA, IHC |
| JAG1 | CSB-PA005638 | JAG1 Antibody | WB, ELISA |
| JAG1 | CSB-PA969766 | JAG1 Antibody | ELISA, IHC |
| JAG1 | CSB-PA234352 | JAG1 Antibody | ELISA, IHC |
| JAG1 | CSB-PA01949A0Rb | JAG1 Antibody | ELISA, IHC, IF |
| JAG1 | CSB-PA01949B0Rb | JAG1 Antibody, HRP conjugated | ELISA |
| JAG1 | CSB-PA01949C0Rb | JAG1 Antibody, FITC conjugated | N/A |
| JAG1 | CSB-PA01949D0Rb | JAG1 Antibody, Biotin conjugated | ELISA |
| Target | Code | Product Name | Detection Range | Sensitivity |
|---|---|---|---|---|
| DLL1 | CSB-EL006947HU | Human Delta-like protein 1(DLL1) ELISA kit | 31.25 pg/ml - 2000 pg/ml | 7.8 pg/ml |
| DLL3 | CSB-EL006948HU | Human Delta-like protein 3(DLL3) ELISA kit | 18.75 pg/mL-1200 pg/mL | 4.68 pg/mL |
| DLL4 | CSB-EL006949HU | Human Delta-like protein 4(DLL4) ELISA kit | 23.5 pg/mL-1500 pg/mL | 5.8 pg/mL |
| DLL4 | CSB-EL006949MO | Mouse Delta-like protein 4(DLL4) ELISA kit | 0.16 ng/ml-10 ng/ml | 0.04ng/ml |
| JAG1 | CSB-EL011927HU | Human Protein jagged-1(JAG1) ELISA kit | 0.156 ng/mL-10 ng/mL | 0.039 ng/mL |
| NOTCH1 | CSB-EL015949HU | Human Neurogenic locus notch homolog protein 1(NOTCH1) ELISA kit | 78 pg/mL-5000 pg/mL | 19.5 pg/mL |
| NOTCH3 | CSB-EL015952HU | Human Neurogenic locus notch homolog protein 3(NOTCH3) ELISA kit | 125 pg/mL-8000 pg/mL | 31.25 pg/mL |
[1] Li X,Yan X,Wang Y, et al. The Notch signaling pathway: a potential target for cancer immunotherapy. J Hematol Oncol. 2023;16 (1):45.
[2] Herranz D,Ambesi-Impiombato A,Palomero T, et al. A NOTCH1-driven MYC enhancer promotes T cell development, transformation and acute lymphoblastic leukemia. Nat Med. 2014;20 (10):1130-7.
[3] Bertulfo K,Perez-Duran P,Miller H, et al. Therapeutic targeting of the NOTCH1 and neddylation pathways in T cell acute lymphoblastic leukemia. Proc Natl Acad Sci U S A. 2025;122 (14):e2426742122.
[4] Lei JH,Xu J,Lyu X, et al. NOTCH1 activation compensates BRCA1 deficiency and promotes triple-negative breast cancer formation. Nat Commun. 2020;11 (1):3256.
[5] Qiu H,Zmina PM,Huang AY, et al. Inhibiting Notch1 enhances immunotherapy efficacy in melanoma by preventing Notch1 dependent immune suppressive properties. Cancer Lett. 2018;434:144-151.
[6] Lindblad KE,Donne R,Liebling I, et al. NOTCH1 Drives Sexually Dimorphic Immune Responses in Hepatocellular Carcinoma. Cancer Discov. 2025;15 (3):495-510.
[7] Lu Y,Cao Y,Guo X, et al. Notch-Targeted Therapeutic in Colorectal Cancer by Notch1 Attenuation Via Tumor Microenvironment-Responsive Cascade DNA Delivery. Adv Healthc Mater. 2024;13 (22):e2400797.
[8] Hubmann R,Schwarzmeier JD,Shehata M, et al. Notch2 is involved in the overexpression of CD23 in B-cell chronic lymphocytic leukemia. Blood. 2002;99 (10):3742-7.
[9] Nayak R,Booker MA,Wang T, et al. Loss of NOTCH2 creates a TRIM28-dependent vulnerability in small cell lung cancer. Dev Cell. 2025;60 (24):3462-3479.e13.
[10] Xu J,Xu W,Yang X, et al. LncRNA MIR99AHG mediated by FOXA1 modulates NOTCH2/Notch signaling pathway to accelerate pancreatic cancer through sponging miR-3129-5p and recruiting ELAVL1. Cancer Cell Int. 2021;21 (1):674.
[11] Lee GH,Yoo KC,An Y, et al. FYN promotes mesenchymal phenotypes of basal type breast cancer cells through STAT5/NOTCH2 signaling node. Oncogene. 2018;37 (14):1857-1868.
[12] Heidari P,Taghizadeh M,Vakili O. Signaling pathways and molecular mechanisms involved in the onset and progression of cerebral autosomal dominant arteriopathy with subcortical infarcts and leukoencephalopathy (CADASIL); a focus on Notch3 signaling. J Headache Pain. 2025;26 (1):96.
[13] Cardinale CJ,Li D,Tian L, et al. Association of a rare NOTCH4 coding variant with systemic sclerosis: a family-based whole exome sequencing study. BMC Musculoskelet Disord. 2016;17 (1):462.
[14] Bonyadi Rad E,Hammerlindl H,Wels C, et al. Notch4 Signaling Induces a Mesenchymal-Epithelial-like Transition in Melanoma Cells to Suppress Malignant Behaviors. Cancer Res. 2016;76 (7):1690-7.
[15] Gramantieri L,Giovannini C,Lanzi A, et al. Aberrant Notch3 and Notch4 expression in human hepatocellular carcinoma. Liver Int. 2007;27 (7):997-1007.
[16] Harb H,Chatila TA. Recent patents in allergy and immunology: Method for treating asthma or allergic disease via anti-Notch4 mAb. Allergy.
[17] Li M,Su B. Up-regulation of NOTCH4 gene expression in bipolar disorder: future studies. Am J Psychiatry. 2013;170 (5):560-1.
[18] Simpson MA,Irving MD,Asilmaz E, et al. Mutations in NOTCH2 cause Hajdu-Cheney syndrome, a disorder of severe and progressive bone loss. Nat Genet. 2011;43 (4):303-5.
[19] McDaniell R,Warthen DM,Sanchez-Lara PA, et al. NOTCH2 mutations cause Alagille syndrome, a heterogeneous disorder of the notch signaling pathway. Am J Hum Genet. 2006;79 (1):169-73.
[20] Brai E,Alina Raio N,Alberi L. Notch1 hallmarks fibrillary depositions in sporadic Alzheimer's disease. Acta Neuropathol Commun. 2016;4 (1):64.
[21] Singh S,Weindorfer C,Nandi A, et al. DLL1-responsive PD-L1 + tumor-associated macrophages promote endocrine resistance in breast cancer. Sci Transl Med. 2025;17 (823):eadr6207.
[22] Xu Y,Qiu Z,Chen J, et al. LINC00460 promotes neuroblastoma tumorigenesis and cisplatin resistance by targeting miR-149-5p/DLL1 axis and activating Notch pathway in vitro and in vivo. Drug Deliv Transl Res. 2024;14 (7):2003-2018.
[23] Xu D,Hu J,Xu S, et al. Dll1/Notch activation accelerates multiple myeloma disease development by promoting CD138+ MM-cell proliferation. Leukemia. 2012;26 (6):1402-5.
[24] Su PL,Chakravarthy K,Furuya N, et al. DLL3-guided therapies in small-cell lung cancer: from antibody-drug conjugate to precision immunotherapy and radioimmunotherapy. Mol Cancer. 2024;23 (1):97.
[25] Rudin CM,Reck M,Johnson ML, et al. Emerging therapies targeting the delta-like ligand 3 (DLL3) in small cell lung cancer. J Hematol Oncol. 2023;16 (1):66.
[26] Owen DH,Giffin MJ,Bailis JM, et al. DLL3: an emerging target in small cell lung cancer. J Hematol Oncol. 2019;12 (1):61.
[27] Hermans BCM,Derks JL,Thunnissen E, et al. DLL3 expression in large cell neuroendocrine carcinoma (LCNEC) and association with molecular subtypes and neuroendocrine profile. Lung Cancer. 2019;138:102-108.
[28] Naseri M,Saeednejad Zanjani L,Vafaei S, et al. Increased cytoplasmic expression of DLL4 is associated with favorable prognosis in colorectal cancer. Future Oncol. 2021;17 (24):3231-3242.
[29] Yan J,Xie Y,Liu Z, et al. DLL4-targeted CAR-T therapy sensitizes neoadjuvant chemotherapy via eliminating cancer stem cells and reshaping immune microenvironment in HER2 + breast cancer. J Immunother Cancer. 2024;12 (11):.
[30] Ding XY,Ding J,Wu K, et al. Cross-talk between endothelial cells and tumor via delta-like ligand 4/Notch/PTEN signaling inhibits lung cancer growth. Oncogene. 2012;31 (23):2899-906.
[31] Wang W,Hu W,Wang Y, et al. Long non-coding RNA UCA1 promotes malignant phenotypes of renal cancer cells by modulating the miR-182-5p/DLL4 axis as a ceRNA. Mol Cancer. 2020;19 (1):18.
[32] Afzalipour R,Abbasi-Dokht T,Sheikh M, et al. The Prediction of DLL4 as a Prognostic Biomarker in Patients with Gastric Cancer Using Anti-DLL4 Nanobody. J Gastrointest Cancer. 2024;55 (3):1380-1387.
[33] Gilbert MA,Keefer-Jacques E,Jadhav T, et al. Functional characterization of 2,832 JAG1 variants supports reclassification for Alagille syndrome and improves guidance for clinical variant interpretation. Am J Hum Genet. 2024;111 (8):1656-1672.
[34] Li G,Liu B,Yang H, et al. Omega-3 polyunsaturated fatty acids alleviate renal fibrosis in chronic kidney disease by reducing macrophage activation and infiltration through the JAG1-NOTCH1/2 pathway. Int Immunopharmacol. 2025;152:114454.
[35] Li C,Wang X,Shi D, et al. RFX5 promotes the progression of triple-negative breast cancer through transcriptional activation of JAG1. Hum Cell. 2025;38 (3):86.
[36] Vaish V,Kim J,Shim M. Jagged-2 (JAG2) enhances tumorigenicity and chemoresistance of colorectal cancer cells. Oncotarget. 2017;8 (32):53262-53275.
[37] Ghoshal P,Nganga AJ,Moran-Giuati J, et al. Loss of the SMRT/NCoR2 corepressor correlates with JAG2 overexpression in multiple myeloma. Cancer Res. 2009;69 (10):4380-7.
[38] Mandula JK,Sierra-Mondragon RA,Chang D, et al. Jagged2 targeting in lung cancer activates anti-tumor immunity via Notch-induced functional reprogramming of tumor-associated macrophages. Immunity. 2024;57 (5):1124-1140.e9.