Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
In multicellular organisms, intercellular connections are essential for maintaining tissue structural integrity and enabling selective transport of substances and signal transduction. Among these, tight junctions (TJs)—specialized intercellular complexes—form critical physiological barriers. The Claudin protein family serves as the core structural component of these tight junctions.
Claudin proteins are widely distributed across various tissues, including the skin, gastrointestinal tract, respiratory system, kidneys, and the blood–brain barrier. They act in concert to ensure that physiological barriers not only effectively block the paracellular penetration of harmful agents (such as pathogens and toxins) but also precisely regulate the selective permeability of water, ions, and small molecules, thereby maintaining internal homeostasis.
Claudin proteins are tetraspanin membrane proteins, with both their N- and C-termini located intracellularly. Their extracellular loops interact with Claudin molecules on adjacent cells to form linear, strand-like structures known as "Claudin strands." These strands tightly fuse the plasma membranes of neighboring cells, sealing the intercellular space and forming the structural backbone of the physical barrier.
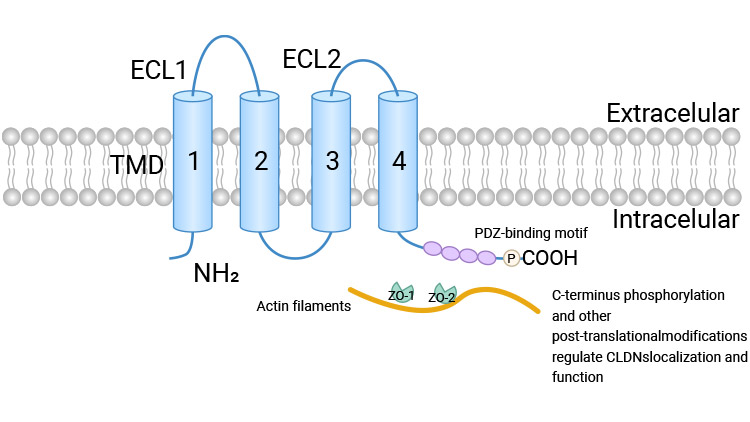
Figure: Schematic Diagram of Claudin Protein Structure [1]
To date, at least 27 Claudin family members (Claudin-1 to Claudin-27) have been identified in the human genome. Despite belonging to the same family, different Claudin proteins exhibit significant differences in tissue distribution, expression levels, and specific functional roles within tight junctions, allowing them to meet the diverse physiological demands of various tissues.
The core functions of Claudin proteins are primarily twofold:
Claudin proteins create continuous intercellular "seals" that effectively prevent the free paracellular diffusion of water-soluble molecules, ions, and microorganisms. This is crucial for maintaining the independence and stability of organ-specific microenvironments—for example, the intestinal barrier blocks harmful substances from entering the systemic circulation, while the blood–brain barrier protects the central nervous system from potentially toxic blood-borne agents.
Beyond their barrier function, specific Claudin proteins can form nanoscale "paracellular aqueous pores" that mediate the selective paracellular transport of ions and small molecules. The identity and combination of Claudin isoforms determine the charge selectivity and size selectivity of these pores, enabling precise reabsorption and secretion of ions such as calcium, magnesium, sodium, and chloride—key processes for maintaining fluid-electrolyte balance and metabolic homeostasis.
Due to their central role in establishing and regulating physiological barriers, dysregulation or aberrant expression of Claudin proteins is closely linked to the pathogenesis of numerous diseases. Consequently, the Claudin family has become a major focus in biomedical research and is widely recognized for its potential as both diagnostic biomarkers and therapeutic targets. Modulating Claudin function offers promising strategies for treating a range of challenging diseases.
Claudin proteins exhibit distinct expression patterns across different tissues and organs. A thorough understanding of these tissue-specific expression profiles is crucial for researchers to select appropriate positive controls, accurately interpret experimental results, and investigate the physiological functions of Claudin proteins.
Table: Expression Profiles of Claudin Proteins in Major Human Tissues and Organs
| Tissue / Organ | Predominantly Expressed Claudin Members | Functional Characteristics |
|---|---|---|
| Skin | CLDN1, CLDN4, CLDN7, CLDN11 | Forms the epidermal barrier to prevent water loss and pathogen invasion |
| Liver | CLDN1, CLDN2, CLDN3, CLDN5 | Maintains hepatocyte polarity and contributes to the bile canalicular barrier |
| Kidney | CLDN2, CLDN4, CLDN7, CLDN8, CLDN10, CLDN16, CLDN19 | Regulates ion-selective reabsorption and barrier function in renal tubules |
| Intestine | CLDN1, CLDN2, CLDN3, CLDN4, CLDN7, CLDN15 | Balances barrier integrity with nutrient/ion paracellular permeability |
| Lung | CLDN1, CLDN3, CLDN4, CLDN5, CLDN18.1 | Maintains airway and alveolar epithelial barrier integrity |
| Stomach | CLDN18.2, CLDN18.1 | Specifically expressed in gastric mucosal epithelium |
| Brain (Blood–Brain Barrier) | CLDN5, CLDN3 | Forms a high-resistance endothelial barrier that restricts substance entry into the brain |
| Testis | CLDN3, CLDN5, CLDN11 | Establishes the blood–testis barrier |
| Mammary Gland | CLDN1, CLDN3, CLDN4, CLDN7 | Maintains ductal epithelial polarity and secretory function |
| Placenta | CLDN1, CLDN4, CLDN5 | Regulates barrier function at the maternal–fetal interface for controlled substance exchange |
Dysregulation of various Claudin family members has been strongly implicated in the pathogenesis and progression of numerous diseases, positioning them as highly valuable biomarkers and potential therapeutic targets. The table below summarizes the associations between specific Claudin members and major diseases, aiming to help researchers rapidly identify relevant targets aligned with their research focus.
Table: Expression of Claudin Family Members Across Human Diseases
| Disease Type | Claudin Member | Expression | Reference |
|---|---|---|---|
|
Breast Cancer
|
CLDN1 | ↓ (down) | [2] |
| CLDN2 | ↑ (up) | [3] | |
| CLDN4 | ↑ | [4] | |
| CLDN7 | ↓ | [5] | |
| CLDN8 | ↑↓ (context-dependent) | [6][7] | |
|
Colorectal cancer
|
CLDN1 | ↑ | [8] |
| CLDN2 | ↑ | [9] | |
| CLDN3 | ↑ | [10] | |
| CLDN4 | ↑ | [9] | |
| CLDN6 | ↓ | [9] | |
| CLDN18 | ↑ | [9] | |
|
Gastric Cancer
|
CLDN1 | ↑ | [11] |
| CLDN3 | ↑ | [12] | |
| CLDN4 | ↑ | [13] | |
| CLDN5 | ↑ | [17] | |
| CLDN6 | ↑ | [14] | |
| CLDN7 | ↑ | [15] | |
| CLDN9 | ↑ | [52] | |
| CLDN18.2 | ↑ | [16] | |
|
Lung cancer(including non-small cell lung
cancer)
|
CLDN1 | ↓ | [18] |
| CLDN2 | ↑ | [19] | |
| CLDN3 | ↓ | [20] | |
| CLDN5 | ↓ | [21] | |
| CLDN6 | ↓ | [22] | |
| CLDN7 | ↓ | [23] | |
|
Hepatocellular carcinoma
|
CLDN1 | ↑ | [24] |
| CLDN3 | ↓ | [25] | |
| CLDN5 | ↓ | [26] | |
| CLDN9 | ↑ | [27] | |
| CLDN11 | ↓ | [28] | |
| CLDN14 | ↓ | [29] | |
| CLDN17 | ↑ | [30] | |
|
Cervical cancer
|
CLDN1 | ↑ | [31] |
| CLDN12 | ↓ | [32] | |
| CLDN18 | ↑ | [33] | |
|
Liver fibrosis
|
CLDN1 | ↑ | [34] |
|
Kidney cancer
|
CLDN3 | ↓ | [35] |
| CLDN4 | ↓ | [36] | |
| CLDN7 | ↓ | [37] | |
| CLDN8 | ↓ | [38] | |
| CLDN10 | ↑ | [39] | |
|
Ovarian cancer
|
CLDN1 | ↓ | [40] |
| CLDN3 | ↑ | [41] | |
| CLDN4 | ↑ | [42] | |
| CLDN5 | ↓ | [43] | |
| CLDN7 | ↑ | [44] | |
|
Thyroid cancer
|
CLDN1 | ↑ | [45] |
|
Pancreatic cancer
|
CLDN1 | ↓ | [46] |
| CLDN4 | ↑ | [47] | |
| CLDN18.2 | ↑ | [48] | |
|
Prostate cancer
|
CLDN1 | ↑ | [49] |
| CLDN3 | ↑ | [50] | |
| CLDN4 | ↑ | [50] | |
| CLDN5 | ↑ | [51] |
The Claudin field is rapidly evolving, with multiple family members emerging as highly promising therapeutic targets and diagnostic biomarkers. Below are several key targets and active research areas currently driving innovation in this space:
CLDN18.2 is a Claudin protein that exhibits
CLDN3 is a core tight junction protein that not only maintains epithelial barrier integrity but also modulates signaling pathways involved in proliferation, metabolism, and immune regulation. Its role in cancer is strikingly tissue-dependent:
This functional duality positions CLDN3 as a high-value but context-sensitive target, requiring careful patient selection for therapeutic development.
Explore CLDN3-related productsCLDN4 exhibits restricted expression in normal tissues but is consistently upregulated across multiple malignancies—including pancreatic, gastric, and ovarian cancers—conferring high tumor selectivity. It plays key roles in regulating cell polarity, signaling pathways, and drug resistance mechanisms, establishing it as a compelling target for immunotherapy. Notably, the CLDN4-targeting bispecific antibody ASP-1002 has entered clinical development and has demonstrated a favorable safety profile with early signs of efficacy, holding promise for addressing unmet needs in refractory solid tumors.
Explore CLDN4-related productsCLDN6 is virtually absent in healthy adult tissues but is re-expressed in multiple cancers—including germ cell tumors, endometrial cancer, and lung cancer—making it an ideal tumor-specific antigen. Its frequent overexpression in immunologically "cold" tumors positions CLDN6 as a promising target to ignite anti-tumor immunity. Cutting-edge modalities such as antibody-drug conjugates (ADCs), CAR-T cell therapies, and bispecific antibodies are advancing rapidly in clinical development, with several pipelines already demonstrating early signs of efficacy.
Explore CLDN6-related productsCLDN9 is a less-studied yet highly promising member of the Claudin family. Recent studies reveal its tumor-specific overexpression in gastric cancer, where it is significantly associated with enhanced tumor invasiveness and poor prognosis. Its restricted expression in normal tissues suggests a favorable therapeutic window. As the mechanistic role of CLDN9 in gastric carcinogenesis becomes clearer, it is emerging as a novel target for differentiated antibody-based therapeutics, offering a potential new avenue for precision treatment of gastric cancer.
Explore CLDN9-related productsCUSABIO offers a complete suite of research tools covering all 27 members of the Claudin family, including high-purity recombinant proteins, antibodies validated across multiple applications (e.g., WB, IHC, FC), and high-sensitivity ELISA kits. Our portfolio supports end-to-end research—from fundamental mechanism studies to translational applications. Whether you are focusing on high-profile targets like CLDN18.2 or CLDN6, or exploring the biological functions of other Claudin members, CUSABIO provides reliable, high-performance solutions to accelerate your discovery.
| Target | Code | Product Name | Source |
|---|---|---|---|
| Cldn1 | CSB-EP005490RA1 | Recombinant Rat Claudin-1 (Cldn1), partial | E.coli |
| CLDN1 | CSB-CF005490HU | Recombinant Human Claudin-1 (CLDN1) | in vitro E.coli expression system |
| CLDN1 | CSB-MP005490HU | Recombinant Human Claudin-1 (CLDN1)-VLPs (Active) | Mammalian cell |
| CLDN1 | CSB-CF005490HUd7 | Recombinant Human Claudin-1 (CLDN1) | in vitro E.coli expression system |
| Cldn1 | CSB-CF005490RA | Recombinant Rat Claudin-1 (Cldn1) | in vitro E.coli expression system |
| CLDN3 | CSB-EP005505HU1a6 | Recombinant Human Claudin-3 (CLDN3), partial | E.coli |
| CLDN3 | CSB-CF005505HU | Recombinant Human Claudin-3 (CLDN3) | in vitro E.coli expression system |
| CLDN3 | CSB-MP005505HU | Recombinant Human Claudin-3 (CLDN3)-VLPs (Active) | Mammalian cell |
| CLDN4 | CSB-MP005506HU | Recombinant Human Claudin-4 (CLDN4)-VLPs (Active) | Mammalian cell |
| CLDN4 | CSB-CF005506HU | Recombinant Human Claudin-4 (CLDN4) | in vitro E.coli expression system |
| CLDN4 | CSB-MP005506HU-D | Recombinant Human Claudin-4 (CLDN4)-Detergent | Mammalian cell |
| Cldn5 | CSB-CF867513RA | Recombinant Rat Claudin-5 (Cldn5) | in vitro E.coli expression system |
| CLDN6 | CSB-CF005508HU | Recombinant Human Claudin-6 (CLDN6), partial | in vitro E.coli expression system |
| CLDN6 | CSB-MP005508HU(A4) | Recombinant Human Claudin-6 (CLDN6)-VLPs (Active) | Mammalian cell |
| CLDN6 | CSB-MP005508HU(A4)f4 | Recombinant Human Claudin-6 (CLDN6), Fluorescent-VLPs (Active) | Mammalian cell |
| CLDN6 | CSB-MP005508HU(A5)-D | Recombinant Human Claudin-6 (CLDN6)-Detergent (Active) | Mammalian cell |
| CLDN6 | CSB-MP6148GKN | Recombinant Chlorocebus sabaeus Claudin (CLDN6)-VLPs (Active) | Mammalian cell |
| Cldn6 | CSB-MP3347MO | Recombinant Mouse Claudin-6 (Cldn6)-VLPs (Active) | Mammalian cell |
| CLDN6 | CSB-BP005508HU(A5) | Recombinant Human Claudin-6 (CLDN6), partial | Baculovirus |
| CLDN6 | CSB-MP005508HU2 | Recombinant Human Claudin-6 (CLDN6), partial-VLPs | Mammalian cell |
| CLDN6 | CSB-MP005508HU1 | Recombinant Human Claudin-6(CLDN6), partial-VLPs | Mammalian cell |
| CLDN6 | CSB-MP005508HU(A4)r2 | Recombinant Human Claudin-6 (CLDN6), Fluorescent-VLPs | Mammalian cell |
| CLDN9 | CSB-MP005511HU | Recombinant Human Claudin-9 (CLDN9)-VLPs (Active) | Mammalian cell |
| CLDN9 | CSB-MP005511HU1-D | Recombinant Human Claudin-9 (CLDN9), partial-Detergent (Active) | Mammalian cell |
| CLDN11 | CSB-CF005492HU | Recombinant Human Claudin-11 (CLDN11) | in vitro E.coli expression system |
| Target | Code | Product Name | Tested Applications |
|---|---|---|---|
| CLDN1 | CSB-PA001656 | CLDN1 Antibody | WB, IHC, IF, ELISA |
| CLDN1 | CSB-PA070223 | CLDN1 Antibody | WB, ELISA |
| CLDN1 | CSB-PA005490GA01HU | CLDN1 Antibody | ELISA |
| CLDN1 | CSB-PA983237 | CLDN1 Antibody | ELISA, WB, IHC |
| CLDN1 | CSB-PA909627 | CLDN1 Antibody | ELISA, WB, IHC |
| CLDN1 | CSB-PA781428 | CLDN1 Antibody | ELISA, WB, IHC |
| CLDN1 | CSB-PA133727 | CLDN1 (Ab-210) Antibody | ELISA, WB |
| CLDN1 | CSB-PA442054 | CLDN1 Antibody | ELISA, WB, IHC |
| CLDN1 | CSB-PA005490LA01HU | CLDN1 Antibody | ELISA, IHC, IF |
| CLDN1 | CSB-PA005490LB01HU | CLDN1 Antibody, HRP conjugated | ELISA |
| CLDN1 | CSB-PA005490LC01HU | CLDN1 Antibody, FITC conjugated | N/A |
| CLDN1 | CSB-PA005490LD01HU | CLDN1 Antibody, Biotin conjugated | ELISA |
| CLDN2 | CSB-PA070222 | CLDN2 Antibody | WB, IHC, ELISA |
| CLDN2 | CSB-PA979246 | CLDN2 Antibody | ELISA, IHC |
| CLDN2 | CSB-PA051134 | CLDN2 Antibody | ELISA, IHC |
| CLDN2 | CSB-PA118188 | CLDN2 Antibody | ELISA, WB, IHC |
| CLDN2 | CSB-PA005500ESR2HU | CLDN2 Antibody | ELISA, IHC, IF |
| CLDN3 | CSB-PA001661 | CLDN3 Antibody | WB, IHC, IF, ELISA |
| CLDN3 | CSB-PA007223 | Phospho-CLDN3 (Y219) Antibody | WB, IHC, ELISA |
| CLDN3 | CSB-PA007225 | CLDN3 Antibody | WB, IHC, ELISA |
| CLDN3 | CSB-PA005505GA01HU | CLDN3 Antibody | ELISA, WB |
| CLDN3 | CSB-PA561954 | CLDN3 Antibody | ELISA, WB, IHC |
| CLDN3 | CSB-PA068230 | CLDN3 Antibody | ELISA, IHC |
| CLDN3 | CSB-PA193301 | CLDN3 Antibody | ELISA, WB, IHC, IF |
| CLDN3 | CSB-PA13119A0Rb | CLDN3 Antibody | ELISA, WB, IHC |
| Target | Code | Product Name | Detection Range | Sensitivity |
|---|---|---|---|---|
| CLDN1 | CSB-EL005490HU | Human Claudin-1(CLDN1) ELISA kit | 15.6 pg/mL-1000 pg/mL | 3.9 pg/mL |
| CLDN2 | CSB-EL005500HU | Human Claudin-2(CLDN2) ELISA kit | 23.5 pg/mL-1500 pg/mL | 5.86 pg/mL |
| CLDN3 | CSB-EL005505HU | Human Claudin-3(CLDN3) ELISA kit | 25 pg/mL-1600 pg/mL | 6.25 pg/mL |
| CLDN4 | CSB-E17961h | Human Claudin 4 (CLDN4)ELISA Kit | 6.25 pg/mL-400 pg/mL | 1.56 pg/mL |
| CLDN5 | CSB-EL005507HU | Human Claudin-5(CLDN5) ELISA kit | 31.25 pg/mL-2000 pg/mL | 7.8 pg/mL |
| CLDN5 | CSB-EL005507MO | Mouse Claudin-5(CLDN5) ELISA kit | 18.75 pg/mL-1200 pg/mL | 4.68 pg/mL |
| CLDN7 | CSB-EL005509HU | Human Claudin-7(CLDN7) ELISA kit | 15.6 pg/mL-1000 pg/mL | 3.9 pg/mL |
[1] Bao L,Yang S,Yang S, et al. Exploring claudin proteins: from sequence motifs to their impact on tight junction-mediated signaling pathways. Amino Acids. 2025;57 (1):48.
[2] Ma F,Zheng S,Ding X, et al. A CLDN1-negative phenotype predicts poor prognosis in triple-negative breast cancer. PLoS One. 2014;9 (11):e112765.
[3] Kimbung S,Kovács A,Fernö M, et al. Claudin-2 is an independent negative prognostic factor in breast cancer and specifically predicts early liver recurrences. Mol Oncol. 2014;8 (1):119-28.
[4] Gao Y,Tang L,Ning K, et al. A novel PAK4-CEBPB-CLDN4 axis involving in breast cancer cell migration and invasion. Biochem Biophys Res Commun. 2019;511 (2):404-408.
[5] West JJ,Golloshi R,Cho CY, et al. Claudin 7 suppresses invasion and metastasis through repression of a smooth muscle actin program. J Cell Biol. 2024;223 (12):.
[6] Lou Y,Jiang WG,Ji W, et al. Knockdown of Claudin-8 (CLDN8) Indicates a Link Between Breast Cancer Cell Sensitivity to Chemotherapeutics and Reveals a Potential Use of CLDN8 as a Molecular Diagnostic and Target for Therapy. Int J Mol Sci. 2025;26 (11):.
[7] Zhang Y,Zheng A,Jin Z, et al. The Expression and Prognostic Significance of Claudin-8 and Androgen Receptor in Breast Cancer. Onco Targets Ther. 2020;13:3437-3448.
[8] Garambois V,Andrade AF,Fauvre A, et al. Improving the response to oxaliplatin by targeting chemotherapy-induced CLDN1 in resistant metastatic colorectal cancer cells. Cell Biosci. 2023;13 (1):72.
[9] Cox KE,Liu S,Hoffman RM, et al. The Expression of the Claudin Family of Proteins in Colorectal Cancer. Biomolecules. 2024;14 (3):.
[10] Andrade-Da-Costa J,De Souza WF,Boroni M, et al. N‑glycosylation and receptor tyrosine kinase signaling affect claudin‑3 levels in colorectal cancer cells. Oncol Rep. 2020;44 (4):1649-1661.
[11] Tannæs TM,Blom GP,Bukholm IR, et al. Up-regulation of CLDN1 in gastric cancer is correlated with reduced survival. BMC Cancer. 2013;13:586.
[12] Danilova NV,Anikina KA,Oleynikova NA, et al. [Claudin-3 expression in gastric cancer]. Arkh Patol. 2020;82 (2):5-11.
[13] Kim WS,Joo MK,Yoo AY, et al. High Expression of Claudin-4 Is Associated with Synchronous Tumors in Patients with Early Gastric Cancer. J Clin Med. 2022;11 (12):.
[14] Yu S,Zhang Y,Zhao G, et al. CLDN6 promotes tumor progression through the YAP1-snail1 axis in gastric cancer. Cell Death Dis. 2019;10 (12):949.
[15] Song Y,Zhao J,Sun J, et al. Claudin-7 (CLDN7) is overexpressed in gastric cancer and promotes gastric cancer cell proliferation, invasion and maintains mesenchymal state. Neoplasma. 2018;65 (3):349-359.
[16] Ruan DY,Luo SX,Dang Q, et al. The antibody-drug conjugate SHR-A1904 for targeting CLDN18.2 in advanced gastric or gastroesophageal junction cancer: a phase 1 trial. Nat Med. 2025;31 (9):3037-3046.
[17] Han L,Huang B,Chen SJ, et al. CLDN5 identified as a biomarker for metastasis and immune infiltration in gastric cancer via pan-cancer analysis. Aging (Albany NY). 2023;15 (11):5032-5051.
[18] Wu YY,Tung CH,Tsai YT, et al. DNA methylation maintains the CLDN1-EPHB6-SLUG axis to enhance chemotherapeutic efficacy and inhibit lung cancer progression. Theranostics. 2020;10 (19):8903-8923.
[19] Yoshino Y,Ikari A. [Cancer Malignancy by Abnormal Claudin Expression]. Gan To Kagaku Ryoho. 2024;51 (11):1100-1104.
[20] Reck M,Hantzsch-Kuhn B,Olchers T, et al. Reduced Claudin-3 Expression Is Linked to Unfavorable Tumor Features and Poor Prognosis in Non-Small Cell Lung Cancer. Lung Cancer (Auckl). 2026;17:1-10.
[21] Kudinov AE,Nikonova AS,Beck TN, et al. Musashi-2 (MSI2) supports TGF-β signaling and inhibits claudins to promote non-small cell lung cancer (NSCLC) metastasis. Proc Natl Acad Sci U S A. 2016;113 (25):6955-60.
[22] Guo H,Li J,Dong Y, et al. CLDN6 inhibited cellular biological function of nonsmall cell lung cancer cells through suppressing aerobic glycolysis via the RIP1/ASK1/JNK axis. J Biochem Mol Toxicol. 2024;38 (3):e23682.
[23] Xia P,Wang W,Bai Y. Claudin-7 suppresses the cytotoxicity of TRAIL-expressing mesenchymal stem cells in H460 human non-small cell lung cancer cells. Apoptosis. 2014;19 (3):491-505.
[24] Muller M,Jühling F,Del Zompo F, et al. Treatment of HCC with claudin-1-specific antibodies suppresses carcinogenic signaling and reprograms the tumor microenvironment. J Hepatol. 2023;78 (2):343-355.
[25] Jiang L,Yang YD,Fu L, et al. CLDN3 inhibits cancer aggressiveness via Wnt-EMT signaling and is a potential prognostic biomarker for hepatocellular carcinoma. Oncotarget. 2014;5 (17):7663-76. doi:10.18632/oncotarget.2288
[26] Sakaguchi T,Suzuki S,Higashi H, et al. Expression of tight junction protein claudin-5 in tumor vessels and sinusoidal endothelium in patients with hepatocellular carcinoma. J Surg Res. 2008;147 (1):123-31.
[27] Liu H,Wang M,Liang N, et al. Claudin-9 enhances the metastatic potential of hepatocytes via Tyk2/Stat3 signaling. Turk J Gastroenterol. 2019;30 (8):722-731.
[28] Yang J,Liu X,Yuan X, et al. miR-99b promotes metastasis of hepatocellular carcinoma through inhibition of claudin 11 expression and may serve as a prognostic marker. Oncol Rep. 2015;34 (3):1415-23.
[29] Li CP,Cai MY,Jiang LJ, et al. CLDN14 is epigenetically silenced by EZH2-mediated H3K27ME3 and is a novel prognostic biomarker in hepatocellular carcinoma. Carcinogenesis. 2016;37 (6):557-566.
[30] Sun L,Feng L,Cui J. Increased expression of claudin-17 promotes a malignant phenotype in hepatocyte via Tyk2/Stat3 signaling and is associated with poor prognosis in patients with hepatocellular carcinoma. Diagn Pathol. 2018;13 (1):72. doi:10.1186/s13000-018-0749-1
[31] Zhang WN,Wang XL,Zhu D, et al. CLDN1 expression in cervical cancer cells is related to tumor invasion and metastasis. Oncotarget. 2016;7 (52):87449-87461.
[32] Kobayashi M,Sugimoto K,Endo Y, et al. Reduced Claudin-12 Expression Predicts Poor Prognosis in Cervical Cancer. Int J Mol Sci. 2021;22 (7).
[33] Yokawa Y,Tanaka T,Mishima R, et al. Claudin-18 expression in gastric type adenocarcinoma and HPV-associated adenocarcinoma of the uterine cervix. Histopathology. 2025.
[34] Saviano A,El Saghire H,Crouchet E, et al. A monoclonal antibody targeting nonjunctional claudin-1 inhibits fibrosis in patient-derived models by modulating cell plasticity. Sci Transl Med. 2022;14 (676):eabj4221.
[35] Büyücek S,Schraps N,Chirico V, et al. Prevalence and clinical significance of Claudin-3 expression in cancer: a tissue microarray study on 14,966 tumor samples. Biomark Res. 2024;12 (1):154.
[36] Owari T,Sasaki T,Fujii K, et al. Role of Nuclear Claudin-4 in Renal Cell Carcinoma. Int J Mol Sci. 2020;21 (21).
[37] Li Y,Gong Y,Ning X, et al. Downregulation of CLDN7 due to promoter hypermethylation is associated with human clear cell renal cell carcinoma progression and poor prognosis. J Exp Clin Cancer Res. 2018;37 (1):276.
[38] Zhu Z,Xu C,Lin L, et al. Prognostic Value and Potential Biological Functions of CLDN8 in Patients with Clear Cell Renal Cell Carcinoma. Onco Targets Ther. 2020;13:9135-9145.
[39] Onagi A,Sugimoto K,Kobayashi M, et al. Extrajunctional CLDN10 cooperates with LAT1 and accelerates clear cell renal cell carcinoma progression. Cell Commun Signal. 2024;22 (1):588.
[40] Zhang H,Lin Y,Kang K, et al. PABPC3 drives ovarian cancer metastasis and drug sensitivity by downregulating CLDN1 expression. Cell Death Dis. 2025;16 (1):840.
[41] He ZY,Wei XW,Luo M, et al. Folate-linked lipoplexes for short hairpin RNA targeting claudin-3 delivery in ovarian cancer xenografts. J Control Release. 2013;172 (3):679-89.
[42] Hu P,Lei L,Wang Y, et al. CLDN4 as a Novel Diagnostic and Prognostic Biomarker and Its Association with Immune Infiltrates in Ovarian Cancer. Mediators Inflamm. 2023;2023:1075265.
[43] Herr D,Sallmann A,Bekes I, et al. VEGF induces ascites in ovarian cancer patients via increasing peritoneal permeability by downregulation of Claudin 5. Gynecol Oncol. 2012;127 (1):210-6.
[44] Dahiya N,Becker KG,Wood WH, et al. Claudin-7 is frequently overexpressed in ovarian cancer and promotes invasion. PLoS One. 2011;6 (7):e22119.
[45] Martínez-Camberos A,Alvarez-Arrazola M,Arámbula-Meraz E, et al. Dysregulation of KRT19, TIMP1, and CLDN1 gene expression is associated with thyroid cancer. Biochem Biophys Res Commun. 2022;617 (Pt 1):55-59.
[46] Zhu L,Tang N,Hang H, et al. Loss of Claudin-1 incurred by DNMT aberration promotes pancreatic cancer progression. Cancer Lett. 2024;586:216611.
[47] Bang C,Park MG,Cho IK, et al. Liposomes targeting the cancer cell-exposed receptor, claudin-4, for pancreatic cancer chemotherapy. Biomater Res. 2023;27 (1):53.
[48] Tojjari A,Idrissi YA,Saeed A. Emerging targets in gastric and pancreatic cancer: Focus on claudin 18.2. Cancer Lett. 2024;611:217362.
[49] Büscheck F,Höflmayer D,Hube-Magg C, et al. Claudin-1 upregulation is associated with favorable tumor features and a reduced risk for biochemical recurrence in ERG-positive prostate cancer. World J Urol. 2020;38 (9):2185-2196.
[50] Maeda T,Murata M,Chiba H, et al. Claudin-4-targeted therapy using Clostridium perfringens enterotoxin for prostate cancer. Prostate. 2012;72 (4):351-60.
[51] Coutinho-Camillo CM,Lourenço SV,da Fonseca FP, et al. Claudin expression is dysregulated in prostate adenocarcinomas but does not correlate with main clinicopathological parameters. Pathology. 2011;43 (2):143-8.
[52] Hu Q,Han L,Wang J, et al. CLDN9 and hsa-miR-4496 as non-invasive biomarkers for gastric cancer detection. Discov Oncol. 2025;16 (1):486.