Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
| Code | CSB-AP004251MO |
| Abbreviation | Recombinant Mouse Il33 protein, partial (Active) |
| MSDS | |
| Size | $204 |
| Order now | |
| Image |
|
| Have Questions? | Leave a Message or Start an on-line Chat |
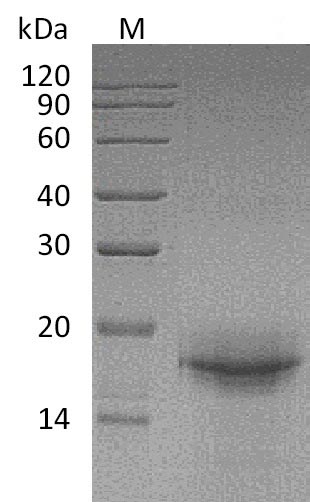
Interleukin-33 drives type 2 immune responses by engaging the ST2 receptor to activate NF-κB and MAPK signaling cascades in ILC2s, mast cells, and Th2 lymphocytes. This tag-free recombinant mouse IL-33 encompasses the mature cytokine domain (aa 109–266), preserving the full receptor-binding region required for faithful ST2-dependent signaling, and demonstrates confirmed binding activity with an ED50 of 0.33 µg/mL for mouse ST2-Fc in functional ELISA. The endotoxin level of less than 1.0 EU/µg minimizes LPS-driven artifacts, which supports use in sensitive cell-based assays such as mast cell activation, ILC2 proliferation, and Th2 differentiation, as well as in vivo murine models of allergic airway inflammation or tissue repair. Purity exceeding 95% by SDS-PAGE, combined with validated ST2 binding, provides a suitable basis for antibody development workflows, ELISA coating antigen applications, and reference standard use in cytokine detection assays.
There are currently no reviews for this product.

