Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
| Code | CSB-EP013065MO |
| Abbreviation | Recombinant Mouse Lpl protein |
| MSDS | |
| Size | US$388 |
| Order now | |
| Image |
|
| Have Questions? | Leave a Message or Start an on-line Chat |
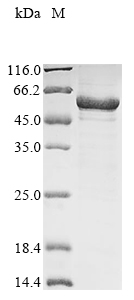
Recombinant Mouse Lipoprotein lipase (Lpl) is produced in E. coli and spans the full length of the mature protein, covering amino acids 28 to 474. The protein carries an N-terminal 10xHis tag and C-terminal Myc tag, which should make purification and detection more straightforward. SDS-PAGE analysis indicates purity levels exceeding 90%, suggesting this product may be suitable for various research applications that require high-quality protein reagents.
Lipoprotein lipase (Lpl) represents a key enzyme in lipid metabolism. It appears to play a central role in breaking down triglycerides found in lipoproteins—chylomicrons and very low-density lipoproteins (VLDL)—into free fatty acids and glycerol. This breakdown process seems essential for maintaining plasma lipid levels and energy balance, which is why Lpl has become an important focus in metabolic and cardiovascular research.
Potential Applications
Note: The applications listed below are based on what we know about this protein's biological functions, published research, and experience from experts in the field. However, we haven't fully tested all of these applications ourselves yet. We'd recommend running some preliminary tests first to make sure they work for your specific research goals.
Based on the provided information, the recombinant Mouse Lipoprotein Lipase (LPL) is expressed in E. coli, a prokaryotic system that is fundamentally unsuitable for producing functional eukaryotic lipases. LPL is a complex enzyme requiring precise folding, glycosylation, dimerization, and interaction with specific cofactors (such as ApoC-II) for its lipolytic activity. E. coli lacks the eukaryotic chaperones, glycosylation machinery, and lipid processing systems necessary for proper LPL folding and activation. The presence of dual tags (N-terminal 10xHis and C-terminal Myc) further increases the risk of improper folding and may sterically hinder functional domains. While the protein is full-length mature (28-474aa) with >90% purity, the expression system makes it highly likely to be misfolded and inactive. Since activity is unverified, the protein cannot be assumed to be correctly folded or bioactive.
1. Antibody Development and Validation Studies
This application is appropriate. The recombinant LPL can serve as an effective immunogen for generating antibodies that recognize linear epitopes, even if the protein is misfolded. The dual tags provide additional epitopes for characterization. However, if LPL is misfolded (as expected in E. coli), antibodies may not recognize conformational epitopes of native, glycosylated LPL in physiological contexts. Validation against native LPL from mammalian sources is essential.
2. Tag-Assisted Protein Interaction Studies
This application is highly problematic. While the dual tags enable technical feasibility for pull-down assays, if LPL is misfolded, it will not interact physiologically with its true binding partners (e.g., lipoproteins, heparin, or ApoC-II). Identified interactions are likely to be non-physiological artifacts. This application should not be pursued without prior validation of proper folding and lipase activity.
3. ELISA Development and Standardization
This application is feasible but with significant limitations. The tags allow development of tag-based detection systems, but if LPL is misfolded, such ELISAs will not measure biologically relevant LPL levels or activity. The assay may detect the recombinant protein, but it won't correlate with functional LPL in biological samples. This approach is only suitable for detecting the recombinant Lipoprotein Lipase itself, not native Lipoprotein Lipase.
4. Biochemical Characterization and Stability Studies
This application is well-suited and should be prioritized. Techniques like circular dichroism spectroscopy, size-exclusion chromatography, and thermal stability assays can directly assess the protein's folding state, oligomerization, and stability. These studies are valuable even if the protein is inactive, as they characterize the recombinant Lipoprotein Lipase itself and can inform about its suitability for other applications.
Final Recommendation & Action Plan
Given the high certainty of misfolding in E. coli for this complex lipase, we recommend first performing comprehensive biophysical characterization (circular dichroism for secondary structure, size-exclusion chromatography with multi-angle light scattering for oligomeric state) to confirm folding status. Antibody development can proceed immediately, but resulting antibodies must be validated against native, glycosylated LPL. Avoid all functional studies (interactions, activity assays) unless proper folding and enzymatic activity are confirmed. For reliable LPL research, obtain the protein from mammalian expression systems capable of proper glycosylation and folding. If using this E. coli-expressed protein, limit applications to antibody production (with validation) and biochemical characterization.
There are currently no reviews for this product.
Do you offer full-length Lipoprotein Lipase (Lpl) recombinant protein from hamster or from mouse? UniProt # P11152
https://www.cusabio.com/Recombinant-Protein/Recombinant-Mouse-Lipoprotein-lipaseLpl-Mammalian-cell-11158383.html
The Lpl recombinant protein should be full-length and without tags at the N- and C-terminus -> Tag-removal service possible
I am interested in the biological activity. Do you offer this kind of service too?
ADAGRDFSDIESKFALRTPEDTAEDTCHLIPGLADSVSNCHFNHSSKTFVVIHGWTVTGMYESWVPKLVAALYKREPDSNVIVVDWLYRAQQHYPVSAGYTKLVGNDVARFINWMEEEFNYPLDNVHLLGYSLGAHAAGVAGSLTNKKVNRITGLDPAGPNFEYAEAPSRLSPDDADFVDVLHTFTRGSPGRSIGIQKPVGHVDIYPNGGTFQPGCNIGEAIRVIAERGLGDVDQLVKCSHERSIHLFIDSLLNEENPSKAYRCNSKEAFEKGLCLSCRKNRCNNLGYEINKVRAKRSSKMYLKTRSQMPYKVFHYQVKIHFSGTEDGKQHNQAFEISLYGTVAESENIPFTLPEVSTNKTYSFLIYTEVDIGELLMMKLKWISDSYFSWPDWWSSPSFVIERIRVKAGETQKKVIFCAREKVSHLQKGKDSAVFVKCHDKSLKKSG

