Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
Histone post-translational modifications are the cornerstone of epigenetic gene regulation, acting by reshaping chromatin architecture and modulating the accessibility of genomic DNA to transcriptional machinery. Early research focused heavily on modifications of the unstructured N-terminal histone tails, which govern transcription primarily by recruiting effector proteins that remodel chromatin states. However, transformative recent discoveries have revealed that modifications within the histone globular domain—especially residues on the DNA-contacting lateral surface—mediate direct, causal control over transcription through structural remodeling of nucleosomes [1].
This article synthesizes classical tail modification mechanisms and cutting-edge research on lateral surface modifications to provide a comprehensive view of how histone marks shape gene expression. We focus on four functionally critical residues, H3K79, H3K122, H3K56, and H3K64, to illustrate distinct regulatory strategies and discuss how dysregulation of these pathways drives human disease.
Table of Contents
3. Pathological Consequences: Dysregulated Histone Modifications Disrupt Gene Expression in Disease
The unstructured N-terminal tails are highly accessible substrates for modifying enzymes, with over 60 identified modification sites including acetylation, methylation, phosphorylation, and ubiquitination [2]. These marks act in combinatorial ways to form a "histone code" that specifies distinct chromatin states and transcriptional outcomes.
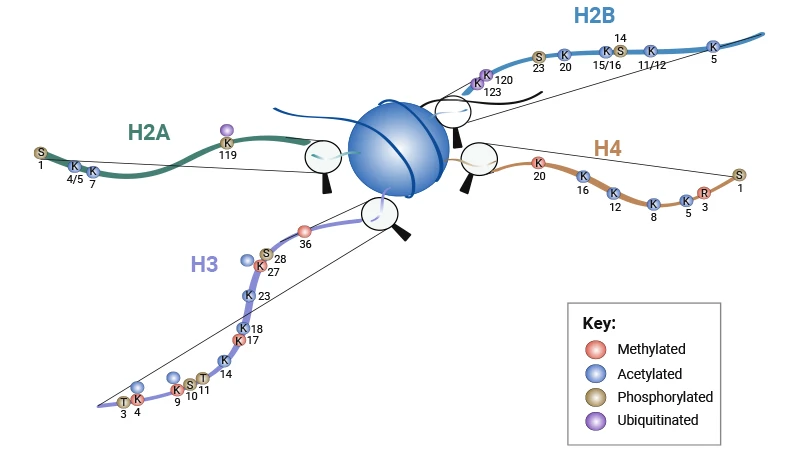
Figure 1. Schematic Showing Histone Tail Modifications [1] (K = lysine, R = arginine, S = serine, T = threonine)
Histone acetylation is universally linked to increased gene expression, acting through two synergistic mechanisms. First, acetyl groups neutralize the positive charge of lysine residues, reducing electrostatic attraction between histones and negatively charged DNA. This loosens nucleosome packing and decondenses higher-order chromatin structure, allowing transcription factors and RNA polymerase to access promoter regions. For example, acetylation of histone H4 at lysine 16 (H4K16ac) directly inhibits the formation of compact 30 nm chromatin fibers and activates transcription both in vitro and in vivo [3].
Second, acetylated lysine residues serve as docking sites for bromodomain-containing co-activators. These effector proteins further recruit chromatin remodelers, additional histone modifiers, and components of the basal transcription machinery, amplifying and sustaining the transcriptional activation signal. In this way, histone acetylation converts a biophysical change in chromatin structure into a robust, coordinated gene expression program.
Unlike acetylation, histone methylation does not alter the charge of modified residues. Instead, it modulates gene expression by recruiting specific effector proteins that either activate or repress transcription, depending on the modified residue and methylation state (mono-, di-, or tri-methylation). Methylated residues are recognized by effector proteins containing chromodomains, Tudor domains, or PHD fingers, which execute downstream regulatory functions.
For instance, trimethylation of histone H3 at lysine 4 (H3K4me3) is enriched at the promoters of actively transcribed genes, where it recruits transcription initiation complexes to drive gene expression. Conversely, H3K27me3 is deposited by Polycomb repressive complexes to silence developmental genes and maintain X-chromosome inactivation [1]. H3K36me3 accumulates along gene bodies to support transcriptional elongation, while H3K9me3 nucleates constitutive heterochromatin to repress repetitive DNA and tissue-inappropriate genes stably.
CUSABIO provides highly validated Histone H3K4me3 Antibody (CSB-PA166887) and Histone H3K27me3 Antibody (CSB-PA963638) for genome-wide mapping of active and repressive chromatin domains with exceptional sensitivity and specificity, enabling precise dissection of gene expression regulatory networks.
Phosphorylation regulates mitotic chromosome condensation (H3S10ph), apoptosis (H2BS14ph), and rapid transcriptional activation [1]. Mono-ubiquitination of H2BK123 is a prerequisite for H3K4 and H3K79 methylation, forming an essential trans-histone regulatory cascade [4]. These modifications add critical layers of temporal and spatial control to gene expression programs.
Phosphorylation and ubiquitination add precise temporal and spatial layers of control over gene expression programs.
Histone phosphorylation occurs on serine and threonine residues, modulating transcription by altering chromatin compaction and recruiting signaling-related effectors. For example, H3S10 phosphorylation drives chromosome condensation during mitosis, which globally pauses transcription; in interphase, rapid phosphorylation of specific histone residues mediates immediate-early gene activation in response to extracellular signals [1]. H2BS14 phosphorylation is linked to apoptotic chromatin condensation and the shutdown of transcriptional activity during programmed cell death.
Histone mono-ubiquitination, particularly of H2B at lysine 123 (H2BK123ub), regulates gene expression through a conserved trans-histone regulatory cascade. This modification is a prerequisite for the methylation of H3K4 and H3K79, two marks tightly associated with active transcription [4]. Through this cascade, H2B ubiquitination coordinates multiple histone modifications to establish and maintain active chromatin states, adding sequential control to gene activation.
Table 1. Histone Tail Modifications [1]
| Histone | Modification | Role |
|---|---|---|
| H2A | H2AS1P | Mitosis; chromath assembly |
| H2AK4/5ac | Transcriptional activation | |
| H2AK7ac | Transcriptional activation | |
| H2AK119P | Spermatogenesis | |
| H2AK119uq | Transcriptional repression | |
| H2B | H2BS14P | Apoptosis |
| H2BS33P | Transcriptional activation | |
| H2BK5ac | Transcriptional activation | |
| H2BK11/12ac | Transcriptional activation | |
| H2BK15/16ac | Transcriptional activation | |
| H2BK20ac | Transcriptional activation | |
| H2BK120uq | Spermatogenesis/meiosis | |
| H2BK123uq | Transcriptional activation | |
| H3 | H3K4me2 | Permissive euchromatin |
| H3K4me3 | Transcriptional elongation; active euchromatin | |
| H3K9me3 | Transcriptional repression; imprinting; DNA methylation | |
| H3R17me | Transcriptional activation | |
| H3K27me3 | Transcriptional silencing; X-inactivation; bivalent genes/gene poising | |
| H3K36me3 | Transcriptional elongation | |
| H3K4ac | Transcriptional activation | |
| H3K9ac | Histone deposition; transcriptional activation | |
| H3K14ac | Transcriptional activation; DNA repair | |
| H1K18ac | Transcriptional activation; DNA repair; DNA replication | |
| H3K23ac | Transcriptional activation; DNA repair | |
| H3K27ac | Transcriptional activation | |
| H3T3P | Mitosis | |
| H3S10P | Mitosis; meiosis; transcriptional activation | |
| H3T11/S28P | Mitosis | |
| H4 | H4R3me | Transcriptional activation |
| H4K20me1 | Transcriptional silencing | |
| H4K20me3 | Heterochromatin | |
| H4K5ac | Histone deposition; transcriptional activation; DNA repair | |
| H4K8ac | Transcriptional activation; DNA repair; transcriptional elongation | |
| H4K12ac | Histone deposition; telomeric silencing; transcriptional activation; DNA repair | |
| H4K16ac | Transcriptional activation; DNA repair | |
| H4S1P | Mitosis |
Unlike Histone tail modifications that act indirectly through effector recruitment, modifications on the histone globular domain control gene expression through a fundamentally different mechanism. The globular domains form the structural core of the nucleosome, and their lateral surface makes extensive, direct physical contacts with wrapped DNA. Modifications in this region often directly alter histone-DNA binding affinity and nucleosome stability, enabling immediate, causal control over transcription without requiring intermediate effector proteins [1].
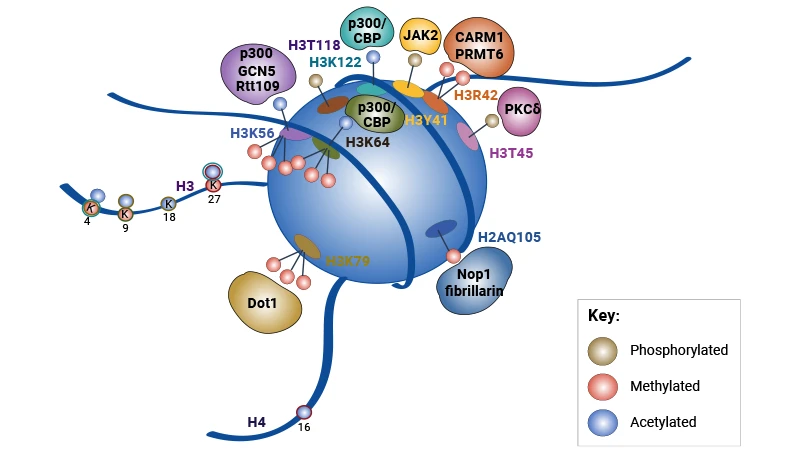
Figure 2. Schematic of a Nucleosome Detailing the Locations of Globular Domain Modifications, the Enzymes
Responsible for Their Deposition, and Their Interplay with Histone Tail Modifications [1]
H3K79 methylation was the first well-characterized globular domain modification, with unique biochemical properties that shape its effects on gene expression. The methyltransferase responsible—DOT1L in mammals, Dot1p in yeast—lacks a canonical SET domain and can only methylate H3K79 on intact, assembled nucleosomes, linking its activity directly to nucleosome structure and ongoing transcription [5,6].
In actively transcribed genomic regions, H3K79me3 is enriched at gene promoters and gene bodies, where it is associated with active transcriptional elongation. In embryonic stem cells, H3K79me1 marks poised bivalent genes that maintain developmental plasticity by keeping genes in a silent but activatable state [7]. Paradoxically, in yeast, H3K79 methylation also contributes to telomeric silencing by preventing the spread of silencing complexes into euchromatic regions, thereby maintaining proper boundaries between active and repressive chromatin domains.
Dysregulation of H3K79 methylation directly disrupts normal gene expression in mixed lineage leukemia (MLL). In MLL-rearranged leukemias, aberrant recruitment of DOT1L to HOX gene loci leads to excessive H3K79 methylation and sustained overexpression of oncogenic HOX genes, driving leukemogenesis [8,9].
CUSABIO's Histone H3K79me3 Antibody (CSB-PA185140) enables high-resolution mapping of DOT1L target genes via ChIP-seq, supporting investigations into how H3K79 methylation dysregulation alters gene expression in hematological malignancies and other diseases.
H3K122 acetylation represents a paradigm shift in understanding how histone modifications control gene expression, as it drives transcription through direct structural disruption of the nucleosome rather than effector recruitment. Located at the nucleosome dyad axis—the position where histone-DNA binding is strongest—acetylation of H3K122 directly weakens these high-affinity interactions, destabilizing the nucleosome and promoting histone eviction from DNA [10].
Critically, unlike tail modifications that are often correlative with active transcription, H3K122ac is sufficient to directly activate transcription in a purified in vitro system, demonstrating a causative rather than merely associative role in gene regulation. In living cells, H3K122ac is deposited within 10 minutes of estrogen-mediated gene induction, 20–30 minutes faster than the canonical active tail mark H3K4me3 [10]. This rapid deposition identifies H3K122ac as an early, direct driver of transcriptional activation, enabling cells to initiate gene expression programs with exceptional speed.
A defining feature of lateral surface modifications is their ability to act as binary switches for gene expression: opposing modifications on the same residue produce diametrically opposite effects on nucleosome stability, enabling rapid transitions between active and repressive chromatin states.
H3K56 is positioned near the DNA entry-exit site of the nucleosome. Acetylation of this residue (H3K56ac) loosens histone-DNA contacts at the nucleosome ends, facilitating DNA unwrapping and promoting transcription, DNA replication, and DNA repair [11,12]. In contrast, trimethylation of H3K56 (H3K56me3) marks repressive heterochromatin and major satellite repeats, stabilizing compact chromatin structure and silencing repetitive DNA elements [13].
Similarly, H3K64 acetylation (H3K64ac) destabilizes nucleosomes and enhances DNA accessibility, thereby activating gene expression. Conversely, H3K64me3 is enriched at pericentromeric heterochromatin, where it silences repetitive elements and maintains genome stability by stabilizing compact chromatin [14,15]. This dual-modification system allows cells to rapidly toggle individual nucleosomes between transcriptionally permissive and restrictive states, providing a flexible mechanism for fine-tuning gene expression.
Table 2. Histone Globular Domain Modifications [1]
| Histone | Site | Modification | Function |
|---|---|---|---|
| H2A | H2AK36 | Acetylation | Enriched at promoters of active genes; relaxes chromatin structure by neutralizing the positive charge of lysine residues, thereby facilitating transcriptional activation. |
| H2AK99 | Methylation | / | |
| H2AQ105 | Methylation | Located on the lateral surface of the nucleosome; no well-defined biological function has been established, and it is speculated to regulate inter-nucleosome interactions. | |
| H2AK119 | Acetylation | Antagonizes the transcriptional repressive effect of H2AK119 ubiquitylation; promotes chromatin opening and participates in transcriptional activation regulation. | |
| H2AK119 | Ubiquitylation | Catalyzed by Polycomb Repressive Complex 1 (PRC1); mediates transcriptional repression, developmental gene silencing, heterochromatin compaction, and X-chromosome inactivation. | |
| H2B | H2BK40 | Methylation | / |
| H2BK82 | Acetylation | Located on the lateral surface of the nucleosome; presumed to promote transcription by weakening electrostatic interactions between histones and DNA, with limited functional studies. | |
| H2BR96 | Methylation | / | |
| H2BK105 | Acetylation | Located near the acidic patch region of the nucleosome; regulates the binding of chromatin regulators to nucleosomes and participates in transcriptional regulation. | |
| H2BK113 | Acetylation | Located in the acidic patch region of the nucleosome; regulates inter-nucleosome interactions and higher-order chromatin structure, and is involved in transcriptional activation. | |
| H2BK117 | Acetylation | Located in the acidic patch region of the nucleosome; regulates chromatin compaction state and participates in the transcriptional activation process. | |
| H3 | H3Y41 | Phosphorylation | Regulates chromatin conformation and nucleosome stability; participates in gene expression regulation and tumorigenesis. |
| H3R42 | Methylation | / | |
| H3T45 | Phosphorylation | Involved in DNA damage response and apoptosis regulation, and mitotic chromosome condensation. | |
| H3R53 | Methylation | / | |
| H3K56 | Acetylation | Regulates nucleosome assembly and stability; facilitates transcriptional activation and DNA damage repair by promoting chromatin disassembly. | |
| H3K56 | Methylation | Competes with acetylation at the same site; has limited functional studies, and is presumed to be involved in transcriptional homeostasis regulation. | |
| H3K64 | Acetylation | Reduces histone-DNA binding affinity at the nucleosome entry-exit site; enhances chromatin accessibility and transcriptional activity. | |
| H3K64 | Methylation | Antagonizes acetylation at the same site; has few functional studies, and is speculated to participate in the maintenance of chromatin state. | |
| H3K79 | Methylation | Catalyzed by DOT1L; associated with active transcriptional elongation, DNA double-strand break repair, and maintenance of genomic stability. | |
| H3K115 | Acetylation | Located in the histone core domain; regulates nucleosome structural stability and participates in transcriptional activation. | |
| H3T118 | Phosphorylation | Regulates nucleosome stability and DNA wrapping strength; promotes chromatin opening and participates in transcriptional regulation. | |
| H3K122 | Acetylation | Disrupts histone octamer-DNA interactions; promotes nucleosome disassembly and strongly enhances transcriptional elongation. | |
| H4 | H4K31 | Acetylation | Enriched at active gene promoter regions; increases chromatin accessibility to support transcriptional activation. |
| H4S47 | Phosphorylation | Participates in DNA damage response; regulates nucleosome stability and the recruitment of repair factors. | |
| H4K59 | Methylation | Preliminary studies suggest it is associated with transcriptional repression, while the specific mechanism and function remain unclear. | |
| H4K77 | Acetylation | Weakens the interaction between histones and DNA; promotes nucleosome sliding and participates in transcriptional regulation. | |
| H4K79 | Acetylation | Regulates nucleosome stability; participates in transcriptional activation and DNA damage repair processes. | |
| H4K91 | Acetylation | Participates in the deposition of newly synthesized histones and chromatin assembly processes. | |
| H4R92 | Methylation | / |
Disruption of histone modification pathways leads to aberrant gene expression programs that underlie numerous human diseases, most notably cancer. Targeting these epigenetic regulators has emerged as a promising therapeutic strategy to restore normal gene expression patterns.
In MLL-rearranged leukemia, DOT1L inhibitors are in advanced clinical trials. By blocking aberrant H3K79 methylation, these agents reverse the pathological overexpression of HOX oncogenes, inducing differentiation and apoptosis in leukemic cells [8,9]. In breast cancer, DOT1L inhibition similarly suppresses the expression of genes driving proliferation, stem cell self-renewal, and metastatic potential, reducing tumor aggressiveness [16].
Histone deacetylase (HDAC) inhibitors, which increase global histone acetylation and reactivate silenced tumor suppressor genes, are already FDA-approved for the treatment of cutaneous T-cell lymphoma, validating the therapeutic potential of targeting histone modification pathways to correct dysregulated gene expression.
Beyond cancer, histone modifications govern stem cell fate and somatic cell reprogramming by reshaping genome-wide gene expression profiles. H3K79 methylation acts as a major epigenetic barrier to reprogramming, and knockdown of DOT1L significantly increases the efficiency of induced pluripotent stem cell (iPSC) generation by removing this block to pluripotency gene expression [17]. Additionally, marks such as H3K56ac and H3K79me2 are critical for the DNA damage response. Their loss leads to genomic instability and aberrant gene expression that promote tumorigenesis [12].
As HDACs represent central therapeutic targets in oncology, neurodegeneration and inflammatory diseases, rigorously validated detection reagents are indispensable for target validation and pharmacodynamic studies of gene expression modulation. CUSABIO provides a full panel of HDAC-specific antibodies and quantitative ELISA kits, all verified for specificity and consistency to support preclinical and translational epigenetic research.
CUSABIO HDAC Research Reagents:
Histone modifications constitute a dynamic, multifaceted system that controls gene expression through two principal mechanistic classes. N-terminal tail modifications primarily act indirectly, recruiting diverse effector proteins to remodel chromatin and assemble or disassemble the transcription apparatus. In contrast, lateral surface modifications on the histone globular domain act directly on nucleosome stability and histone-DNA interactions, providing a more immediate mechanism for transcriptional control.
Despite major advances, key questions remain. The complex crosstalk between tail and globular domain modifications in coordinating gene expression programs is not fully understood, and the identity of specific "reader" proteins for many globular domain marks remains elusive. Addressing these questions will be critical to fully unraveling the epigenetic logic of gene regulation.
As epigenetic research advances, CUSABIO remains committed to developing innovative, high-quality reagents that enable breakthrough discoveries. Our comprehensive epigenetic toolkit empowers researchers worldwide to unravel the complexities of histone-mediated gene regulation and translate these findings into novel therapeutic strategies. Visit https://www.cusabio.com/ to explore our full product catalog and request a free sample of our most popular histone modification antibodies.
Epigenetic Histone Modifications Pathway
How to Measure Histone Modification?
References
[1] Lawrence M, Daujat S, Schneider R. Lateral Thinking: How Histone Modifications Regulate Gene Expression [J]. Trends Genet. 2016; 32(1): 42-56.
[2] Kouzarides T. Chromatin modifications and their function [J]. Cell. 2007; 128(4): 693-705.
[3] Akhtar, A., & Becker, P. B. (2000). Activation of Transcription through Histone H4 Acetylation by MOF, an Acetyltransferase Essential for Dosage Compensation in Drosophila [J]. Molecular Cell, 5(2), 367-375.
[4] Briggs SD, Xiao T, Sun ZW, et al. Gene silencing: trans-histone regulatory pathway in chromatin [J]. Nature. 2002; 418(6897): 498.
[5] van Leeuwen F, Gafken PR, Gottschling DE. Dot1p modulates silencing in yeast by methylation of the nucleosome core [J]. Cell. 2002; 109(6): 745-756.
[6] Feng Q, Wang H, Ng HH, et al. Methylation of H3-lysine 79 is mediated by a new family of HMTases without a SET domain [J]. Curr Biol. 2002; 12(12): 1052-1058.
[7] Steger DJ, Lefterova MI, Ying L, et al. DOT1L/KMT4 recruitment and H3K79 methylation are ubiquitously coupled with gene transcription in mammalian cells [J]. Mol Cell Biol. 2008; 28(8): 2825-2839.
[8] Okada Y, Feng Q, Lin Y, et al. hDOT1L links histone methylation to leukemogenesis [J]. Cell. 2005; 121(2): 167-178.
[9] Okada Y, Jiang Q, Lemieux M, et al. Leukaemic transformation by CALM-AF10 involves upregulation of Hoxa5 by hDOT1L [J]. Nat Cell Biol. 2006; 8(9): 1017-1024.
[10] Tropberger P, Pott S, Keller C, et al. Regulation of transcription through acetylation of H3K122 on the lateral surface of the histone octamer [J]. Cell. 2013; 152(4): 859-872.
[11] Neumann H, Hancock SM, et al. A method for genetically installing site-specific acetylation in recombinant histones defines the effects of H3 K56 acetylation [J]. Mol Cell. 2009 Oct 9;36(1):153-63.
[12] Masumoto H, Hawke D, Kobayashi R, Verreault A. A role for cell-cycle-regulated histone H3 lysine 56 acetylation in the DNA damage response [J]. Nature. 2005; 436(7048): 294-298.
[13] Jack AP, Kim J, Kim JH, et al. H3K56me3 is a novel, conserved heterochromatic mark that largely but not completely overlaps with H3K9me3 in both regulation and localization [J]. PLoS ONE. 2013; 8(2): e51765.
[14] Di Cerbo V, Mohn F, Ryan DP, et al. Acetylation of histone H3 at lysine 64 regulates nucleosome dynamics and facilitates transcription [J]. Elife. 2014; 3: e01632.
[15] Daujat S, Weiss T, Mohn F, et al. H3K64 trimethylation marks heterochromatin and is dynamically remodeled during developmental reprogramming [J]. Nat Struct Mol Biol. 2009; 16(7): 777-781.
[16] Zhang L, Chen X, Li Y, et al. Inhibition of histone H3K79 methylation selectively inhibits proliferation, self-renewal and metastatic potential of breast cancer [J]. Oncotarget. 2014; 5(21): 10665-10677.
[17] Onder TT, Karaoz U, Murphy N, et al. Chromatin-modifying enzymes as modulators of reprogramming [J]. Nature. 2012; 483(7391): 598-602.


Comments
Leave a Comment