Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
B7-H3 (CD276), the "dual-faced immune sentinel," reached a watershed moment in 2025. In January 2025, Roche committed an upfront payment of $570 million to secure YL201, an ADC drug from Yilian Bio. That December, its Phase I data were published in Nature Medicine, demonstrating a 63.9% objective response rate in extensive-stage small cell lung cancer—surpassing the historical benchmark set by topotecan. Almost simultaneously, Merck & Co./Daiichi Sankyo’s I-DXd had its Phase III clinical hold lifted by the FDA by the end of 2025, while BioNTech announced plans to initiate a pivotal Phase III trial of its B7-H3 ADC in first-line prostate cancer in 2026. Domestically, the CAR-T therapy MT027 received FDA approval in December 2025 to enter Phase II trials for glioblastoma. The first B7-H3-targeted therapy is expected to launch globally in 2027, with the market projected to start at $300 million and grow at a compound annual growth rate of 74%, reaching $5.5 billion by 2032. Chinese innovators already hold three late-stage "tickets," positioning this once-debated laboratory molecule firmly on China’s fast-track timeline for immuno-oncology 2.0. This article will systematically review the structure, function, and tumor association of B7-H3, integrate the latest advances outlined above, and explore its future therapeutic directions.
In cellular immune response, the proliferation and activation of T cells requires not only the first signal presented by APC or MHC on the surface of tumor cells, but also the second signal provided by costimulatory molecules [1]. The B7-CD28 superfamily is one of the newly discovered costimulatory molecules and belongs to the immunoglobulin superfamily.
The B7 family of molecules can provide stimulating signals to enhance and maintain the T cell immune response, or produce inhibitory signals to limit and weaken the T cell immune response. Therefore, the family plays an important role in tumor diseases, organ transplantation and autoimmune diseases.
According to the functions, the B7 family can be divided into three groups [2].
Group 1: B7-1 (CD80) and B7-2 (CD86).
Group 2: B7-H1 (PD-L1) and B7-DC (PD-L2)
Group 3: B7-H3 (CD276) and B7-H4 (B7x). Their receptors have not been identified, but are thought to be involved in co-stimulation and co-inhibition pathways.
Table 1 List of B7 family members
| B7 molecule | Alternate name | Receptor | Function |
|---|---|---|---|
| B7-1 | CD80 | CD28, CTLA-4 | Stimulatory |
| B7-2 | CD86 | CD28, CTLA-4 | Stimulatory |
| B7-H2 | ICOS-L | ICOS, CD28, CTLA-4 | Stimulatory |
| B7-H1 | PD-L1, CD274 | PD-1, CD80 | Inhibitory |
| B7-DC | PD-L2, CD273 | PD-1 | Inhibitory |
| B7-H3 | CD276 | TLT-2 (Mouse) | Stimulatory / Inhibitory |
| B7-H4 | B7x, B7S1 | / | Stimulatory / Inhibitory |
B7 homolog 3 protein (B7-H3), also known as CD276, is an important immune checkpoint molecule in the B7-CD28 family. It is a member of the B7 family identified in the cDNA library of human dendritic cells in 2001 [3]. CD276 is mainly present in membrane proteins and soluble forms. Soluble CD276 (soluble CD276, sCD276) is cleaved by membrane proteins by metalloproteinases [4].
In addition, B7-H3 protein was also found in exosomes and other extracellular vesicles. CD276 is a T cell co-suppressor molecule with partial costimulatory function. CD276 can effectively inhibit the function of T cells and NK cells, and also has an effect on bone development.
CD276 is expressed in a variety of malignant tumors, and is closely related to the growth, metastasis, recurrence and poor prognosis of malignant tumors.
CD276 is a type I transmembrane glycoprotein consisting of 316 amino acids with a molecular weight of 45-66 kDa, which has a similar molecular structure to B7-H1 (PD-L1).
It contains a putative 28AA signal peptide, an extracellular region of 217AA consisting of immunoglobulin constant (IgC) and variable (IgV) structures, a transmembrane region and a 45 amino acid cytoplasmic domain. CD276 gene consists of 10 exons, of which 4-7 exons encode the extracellular IgV-IgC domain.
B7-H3 has been found to exist in two forms: 2Ig-B7-H3 and 4Ig-B7-H3. 2Ig-B7-H3 is expressed in murine and human cells with extracellular IgV-IgC structure; 4Ig-B7-H3 is expressed only in human cells and consists of a tandem repeat of the IgV-IgC-IgV-IgC structure. The predominant form of human B7-H3 is 4IgB7-H3, located on chromosome 15. The mouse B7-H3 gene is located on chromosome 9 [5]. B7-H3 is one of the most evolutionarily conservative members of the B7 family, as it is commonly expressed in species ranging from bony fish to mammals.
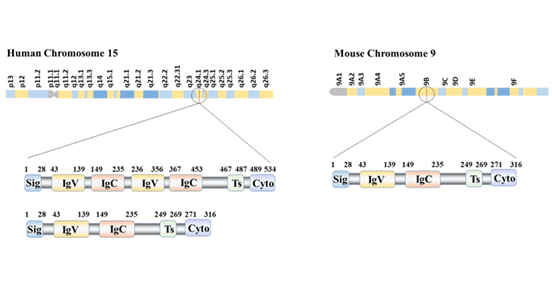
Figure 1 B7-H3 structure and isoforms
CD276 transcripts are widely expressed in tissues such as heart, liver, placenta, prostate, testis, uterus, pancreas, small intestine, and colon. The expression of the CD276 protein is more restricted to the cell surface, such as activated dendritic cells, monocytes, T cells, B cells, and NK cells [6].
Abnormally high expression of CD276 in a variety of cancer cells or tissues, including gastric cancer, lung cancer, prostate cancer, kidney cancer, pancreatic cancer, ovarian cancer, breast cancer, endometrial cancer, liver cancer, colorectal cancer, oral cancer, bladder cancer, osteosarcoma and hematologic malignancies.
The molecular mechanism regulating the expression of CD276 is unclear, but the expression of CD276 protein is inversely proportional to the level of miR-29, and the miR-29 binding site on CD276 is evolutionarily conserved.
The receptor for human B7-H3 has not been found. However, in mouse models, TLT-2, a trigger receptor expressed in the myeloid cell (TREM) receptor family, has been identified as a potential receptor for mouse B7-H3 [7].
TLT-2 is a type I transmembrane protein with a single IgV domain, a transmembrane region and a short cytoplasmic tail [8] [9]. The IgV domain has two potential N-glycosylation sites, a serine/threonine rich region and several predicted O-linked glycosylation sites.
TLT-2 has been shown to be a costimulatory TCR for B7-H3. The interaction of B7-H3 with TLT-2 will result in an increase in IL-2 and IFN-γ production. Blocking the interaction between B7-H3 and TLT-2 with a mAb against B7-H3 or TLT-2 is effective in inhibiting CD8+ T cell-mediated contact hypersensitivity [7].
T cells: B7-H3 may play a dual role in the immune system. On the one hand, B7-H3, as a costimulatory molecule, can co-stimulate CD4+ and CD8+ cells, induce cellular immunity, and selectively enhance the production of interferon-gamma (IFNG) in T-cell receptor signaling.
On the other hand, experiments in humans and mice have demonstrated that B7-H3 also has a co-inhibitory effect [10], which can inhibit Treg cells, thereby allowing tumor escape from the immune response. This may be related to NFAT, NF-kappa B and AP-1 factors [11]. B7-H3 indirectly activated NF-κΒ, PI3K/Akt, JAK/STAT3 pathway induced cell survival and proliferation. B7-H3 also increases ROS and HIF1a levels, leading to increased glycolysis.
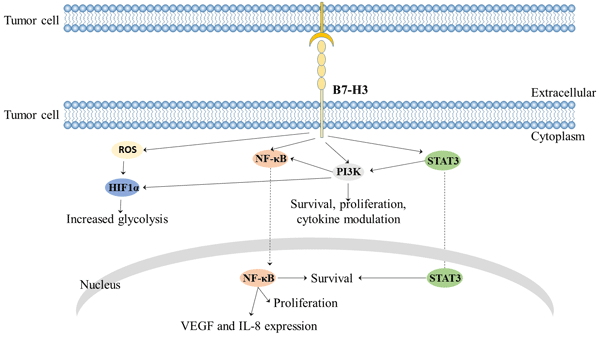
Figure 2 B7-H3 mediated signaling pathway
NK cells: CD276 inhibits NK cell activity in tissue culture, thereby reducing the function of NK cells. This suggests that there should be a receptor for B7-H3 on NK cells.
In gliomas, a higher tumor grade is associated with a higher percentage of CD276 expression. Both soluble CD276 and cell-bound CD276 are capable of inhibiting NK cell-mediated cleavage.
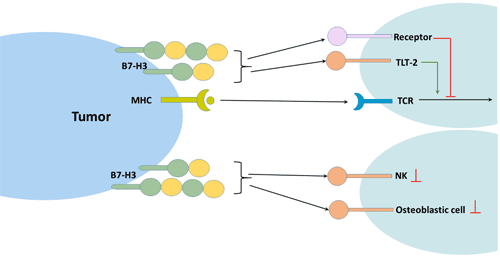
Figure 3 B7-H3 and immune response
B7-H3 plays an important role in tumor immune escape by inhibiting T cells. In addition, it plays an important role in the non-immune system. Studies have shown that B7-H3 is involved in the regulation of cancer cell metastatic potential, and may also affect the biological characteristics of tumor colonization and invasion.
High expression of B7-H3 in cell lines of bladder adenocarcinoma, prostate cancer and melanoma has been shown to promote invasion and metastasis of tumor cells. In melanoma cells, silencing B7-H3 can reduce the invasion ability of matrix gel. This may be because B7-H3 silencing reduces the expression of transfer-related proteins, such as matrix metalloproteinase (MMP)-2, signal transduction and transcriptional activator 3 (STAT3), and IL-8.
B7-H3 molecules are highly expressed in lung cancer, colorectal cancer, membrane adenocarcinoma and other cancer tissues, and are associated with tumor spread or poor prognosis.
B7-H3 is expressed in a variety of different non-small cell lung cancer cell lines [12]. Studies in patients with NSCLC have shown that B7-H3 has an inhibitory effect on the immune system of NSCLC [13].
High expression of B7-H3 protein is associated with poor prognosis [14], and B7-H3 can also be involved in the development of NSCLC by inducing monocytes to develop into anti-inflammatory cells [15].
The expression between B7-H1 (CD80) and B7-H3 was positively correlated, and their expression was significantly correlated with NSCLC lymph node metastasis. B7-H3 may be a potential marker for the diagnosis of NSCLC invasion and metastasis, but it also requires a large amount of clinical data to prove it.
B7-H3 is abnormally expressed in prostate cancer, which may play a role in regulating the proliferation and invasion of the prostate cancer cell line PC-3 [16]. In addition, B7-H3 can also inhibit the immune response by regulating Treg cells, thus mediating the tumor escape immune response. Higher B7-H3 expression was associated with an increased risk of biochemical recurrence after radiotherapy [17].
In addition, B7-H3 is highly expressed in renal cell carcinoma tumor tissues and perivascular tumors, and miR-187 overexpression in renal cell carcinoma may inhibit tumor proliferation and metastasis by reducing the mRNA level of B7-H3 [18].
In breast cancer patients, miR-29c is effective in inhibiting B7-H3 expression [19].
In a mouse model of breast cancer, it was found that B7-H3 promotes the stabilization of hypoxia-inducible factor-1 (HIF1α) through reactive oxygen species, thereby promoting tumor uptake of glucose and growth [20].
The expression of B7-H3 in primary tumors is significantly associated with tumor size and lymphatic invasion. B7-H3 is a potential predictor of future diagnosis of breast cancer [21].
B7-H3 has been shown to be expressed in most malignant neuroectodermal tumors, and B7-H3 expression levels are positively correlated with histological tumor grade [22].
In hepatocellular carcinoma tissues, B7-H3 expression was significantly enhanced, and was associated with increased risk of recurrence, poor pathological type, and poor prognosis. Its mechanism of action may be that B7-H3 causes tumors to evade T cell-associated immune responses. The STAT3 signaling pathway mediated by B7-H3 may accelerate the progression of hepatocellular carcinoma by inducing M2-type polarization of tumor-associated macrophages. B7-H3 is abnormally elevated in the blood of patients with hepatocellular carcinoma [23], demonstrating its important role in the early diagnosis and prognosis of hepatocellular carcinoma.
B7-H3 is highly expressed in various tumor tissues and is closely related to tumor progression. The tail of B7-H3 cytoplasm is short, has no known signal motif, and its receptor has not been recognized. Nevertheless, B7-H3 molecule is considered as a promising immunotherapy research direction.
Currently, most targeted therapies regard B7-H3 as a negative regulator of T-cell-mediated immune response in tumors, and specific blocking of B7-H3 could provide a new targeted therapy similar to anti-CTLA-4 mAb therapy.
Human cancer immunotherapy strategies for B7-H3 mainly include the following:
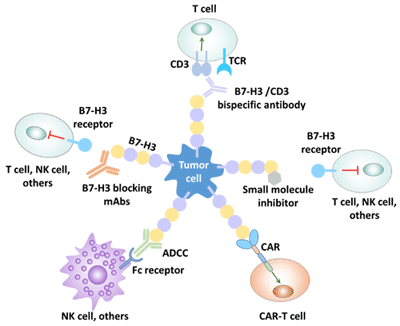
Figure 4 Human immunotherapy strategy for B7-H3
Enoblituzumab (MGA271), which is being studied, is a monoclonal antibody (mAb) of B7-H3 that exerts antitumor activity by antibody-dependent cell-mediated cytotoxicity (ADCC). In a clinical phase I trial (test NCT01391143), the antibody was shown to have good tolerance and antitumor activity.
An antibody drug bound to 8H9, a monoclonal antibody labeled iodine-131, has been clinically successful in the rescue and treatment of patients with metastatic central nervous system tumors of neuroblastoma. Phase I CD276 antibody clinical trials of radiolabolic marker 8H9 in patients with peritoneal cancer, glioma and advanced central nervous system cancer (NCT01099644, NCT01502917 and NCT00089245) are currently under way.
Anti-B7-H3 mAb MJ18 inhibited tumor growth in a pancreatic cancer model.
The combination of B7-H3 mAb and chemotherapy was a preclinical success. By blocking the expression of B7-H3 in combination with anti-tumor drugs, the therapeutic effect was better than that of anti-tumor drugs alone [24].
B7-H3 CAR-T therapy for glioblastoma is currently in Phase Ⅱ.
B7-H3 protein
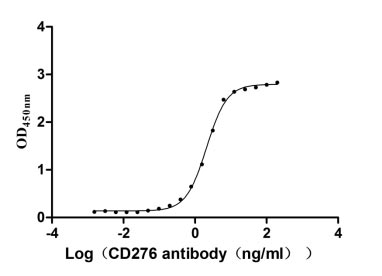
Recombinant Human CD276 antigen(CD276),partial (Active) (Code: CSB-MP733578HU)
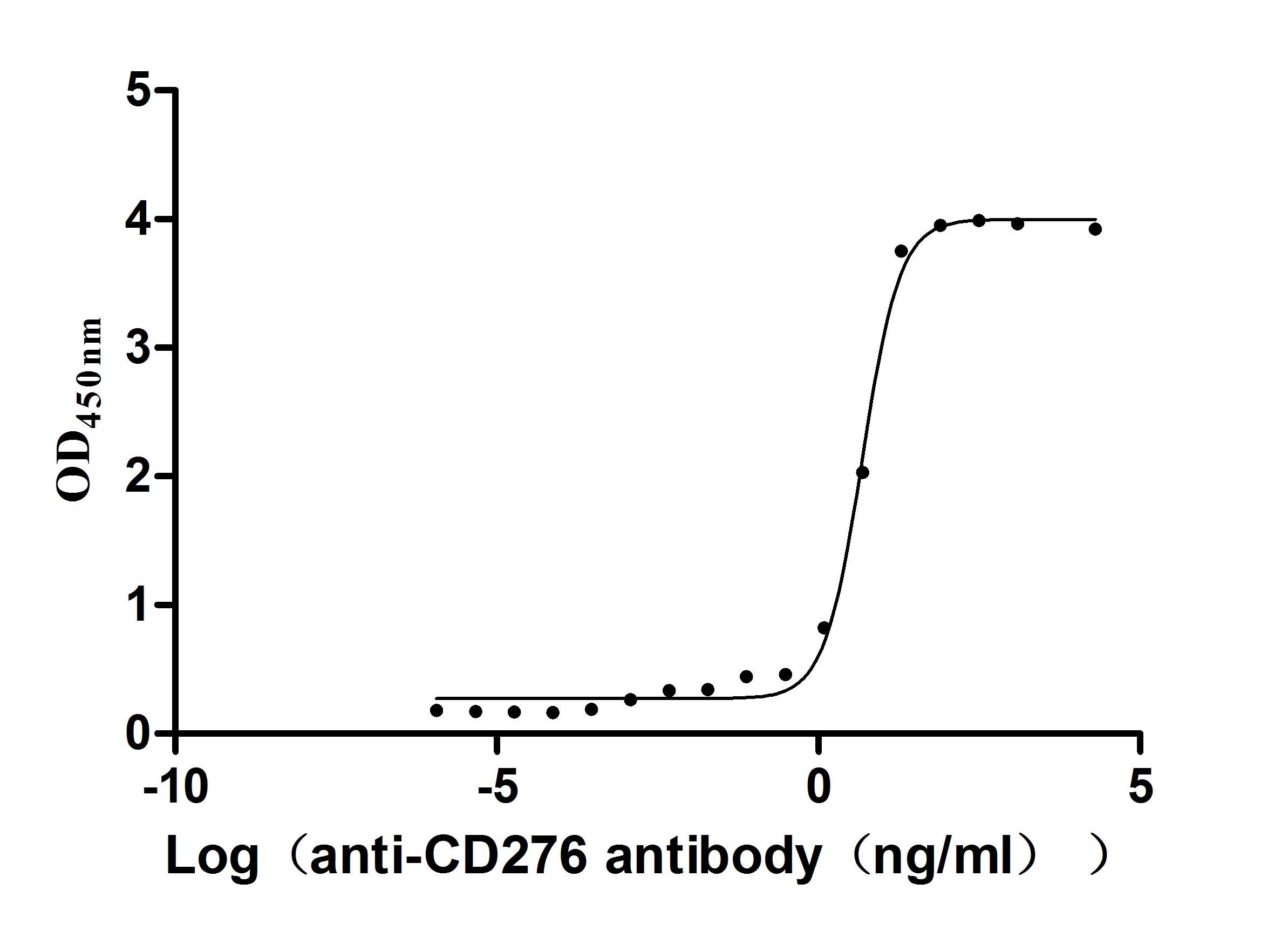
Recombinant Macaca fascicularis CD276 molecule(CD276),partial (Active) (Code: CSB-MP5140MOV)
References
[1] Hodge J W, Greiner J W, Tsang K Y, et al. Costimulatory molecules as adjuvants for immunotherapy [J]. Front Biosci, 2006, 11(788-803).
[2] Seliger B, Marincola F M, Ferrone S, et al. The complex role of B7 molecules in tumor immunology [J]. Trends in molecular medicine, 2008, 14(12): 550-559.
[3] Chapoval A I, Ni J, Lau J S, et al. B7-H3: a costimulatory molecule for T cell activation and IFN-γ production [J]. Nature immunology, 2001, 2(3): 269.
[4] Zhang G, Hou J, Shi J, et al. Soluble CD276 (B7-H3) is released from monocytes, dendritic cells and activated T cells and is detectable in normal human serum [J]. Immunology, 2008, 123(4): 538-546.
[5] Sun M, Richards S, Prasad D V R, et al. Characterization of mouse and human B7-H3 genes [J]. The Journal of Immunology, 2002, 168(12): 6294-6297.
[6] Wang L, Kang F B, Shan B E. B7-H3-mediated tumor immunology: Friend or foe? [J]. International journal of cancer, 2014, 134(12): 2764-2771.
[7] Hashiguchi M, Kobori H, Ritprajak P, et al. Triggering receptor expressed on myeloid cell-like transcript 2 (TLT-2) is a counter-receptor for B7-H3 and enhances T cell responses [J]. Proceedings of the National Academy of Sciences, 2008, 105(30): 10495-10500.
[8] King R G, Herrin B R, Justement L B. Trem-like transcript 2 is expressed on cells of the myeloid/granuloid and B lymphoid lineage and is up-regulated in response to inflammation [J]. The Journal of Immunology, 2006, 176(10): 6012-6021.
[9] Allcock R J N, Barrow A D, Forbes S, et al. The human TREM gene cluster at 6p21. 1 encodes both activating and inhibitory single IgV domain receptors and includes NKp44 [J]. European journal of immunology, 2003, 33(2): 567-577.
[10] Veenstra R G, Flynn R, Kreymborg K, et al. B7-H3 expression in donor T cells and host cells negatively regulates acute graft-versus-host disease lethality [J]. Blood, 2015, 125(21): 3335-3346.
[11] Hofmeyer K A, Ray A, Zang X. The contrasting role of B7-H3 [J]. Proceedings of the National Academy of Sciences, 2008, 105(30): 10277-10278.
[12] Boland J M, Kwon E D, Harrington S M, et al. Tumor B7-H1 and B7-H3 expression in squamous cell carcinoma of the lung [J]. Clinical lung cancer, 2013, 14(2): 157-163.
[13] Mao Y, Li W, Chen K, et al. B7-H1 and B7-H3 are independent predictors of poor prognosis in patients with non-small cell lung cancer [J]. Oncotarget, 2015, 6(5): 3452.
[14] Chen L, Zhang G, Sheng S, et al. Upregulation of soluble B7-H3 in NSCLC-derived malignant pleural effusion: A potential diagnostic biomarker correlated with NSCLC staging [J]. Clinica Chimica Acta, 2016, 457: 81-85.
[15] Sun J, Mao Y, Zhang Y Q, et al. Clinical significance of the induction of macrophage differentiation by the costimulatory molecule B7-H3cancer [J]. Oncology letters, 2013, 6(5): 1253-1260.
[16] Yuan H, Wei X, Zhang G, et al. B7-H3 over expression in prostate cancer promotes tumor cell progression [J]. The Journal of urology, 2011, 186(3): 1093-1099.
[17] Parker A S, Heckman M G, Sheinin Y, et al. Evaluation of B7-H3 expression as a biomarker of biochemical recurrence after salvage radiation therapy for recurrent prostate cancer [J]. International Journal of Radiation Oncology Biology Physics, 2011, 79(5): 1343-1349.
[18] Zhao J, Lei T, Xu C, et al. MicroRNA-187, down-regulated in clear cell renal cell carcinoma and associated with lower survival, inhibits cell growth and migration though targeting B7-H3 [J]. Biochemical and biophysical research communications, 2013, 438(2): 439-444.
[19] Nygren M K, Tekle C, Ingebrigtsen V A, et al. Identifying microRNAs regulating B7-H3 in breast cancer: the clinical impact of microRNA-29c [J]. British journal of cancer, 2014, 110(8): 2072.
[20] Lim S, Liu H, Da Silva L M, et al. Immunoregulatory protein B7-H3 reprograms glucose metabolism in cancer cells by ROS-mediated stabilization of HIF1α [J]. Cancer research, 2016, 76(8): 2231-2242.
[21] Wilson K E, Bachawal S V, Abou-Elkacem L, et al. Spectroscopic photoacoustic molecular imaging of breast cancer using a B7-H3-targeted ICG contrast agent [J]. Theranostics, 2017, 7(6): 1463.
[22] Zhou Z, Luther N, Ibrahim G M, et al. B7-H3, a potential therapeutic target, is expressed in diffuse intrinsic pontine glioma [J]. Journal of neuro-oncology, 2013, 111(3): 257-264.
[23] Zhao L, Xie C, Liu D, et al. Early detection of hepatocellular carcinoma in patients with hepatocirrhosis by soluble B7-H3 [J]. Journal of Gastrointestinal Surgery, 2017, 21(5): 807-812.
[24] Zhang W, Wang J, Wang Y, et al. B7-H3 silencing by RNAi inhibits tumor progression and enhances chemosensitivity in U937 cells [J]. 2015.


Comments
Leave a Comment