Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
The human genome comprises between 20,000 and 25,000 protein-encoding genes [1], while the human proteome is estimated at up to 1,000,000 proteins [2]. Why is the human proteome size far larger than the human gene set? In addition to alternative splicing of mRNA as a source of protein diversity, post-translational modifications (PTMs) of proteins further affect and increase the spectrum of possible protein variability and complexity.
According to the "central dogma" of biology, the DNA is first transcribed to RNA and then translated to proteins. However, most proteins undergo some additional steps after their biosynthesis, which are necessary for cells, tissues, and organisms to achieve their functional biology and diversity. These enzymatic modifications of proteins following protein translation are known as PTMs.
1. What Are Post-translational Modifications?
2. Classifications of Post-translational Modifications
3. The biological Significance of Post-translational Modifications
Following biosynthesis, proteins are further tailored in a variety of ways to complete their structure, designate their spatial orientation, or regulate their activity. These dynamic, tightly orchestrated processing events that alter the characteristics of a protein via proteolytic cleavage and the addition of functional groups (e.g. acetyl, phosphoryl, and glycosyl) to a single type of amino acid or multiple amino acids are known as post-translational modifications (PTMs) [3]. PTMs are frequently found in secretory proteins, membrane proteins, histones, and other proteins with important structures/functions. They occur in the nucleus, cytoplasm, endoplasmic reticulum, and Golgi apparatus, among other cellular organelles [4]. And PTMs are either reversible reactions like covalent modifications or irreversible reactions such as proteolytic modifications.
It is estimated that enzymes that perform various PTMs account for 5% of the proteins of the entire proteome. These enzymes include kinases, phosphatases, transferases, and ligases, which add or remove functional groups, proteins, lipids, or sugars to or from amino acid side chains, as well as proteases, which remove specific sequences or regulatory subunits from amino acid side chains.
More than 400 different forms of PTMs have been identified. They can be classified according to the type of amino acid side chain modified, the category of the modifying enzyme, and the extent of reversibility. The most studied PTMs include phosphorylation, acetylation, ubiquitination, methylation, glycosylation, SUMOylation, lipidation (palmitoylation, myristoylation, and prenylation), and sulfation.
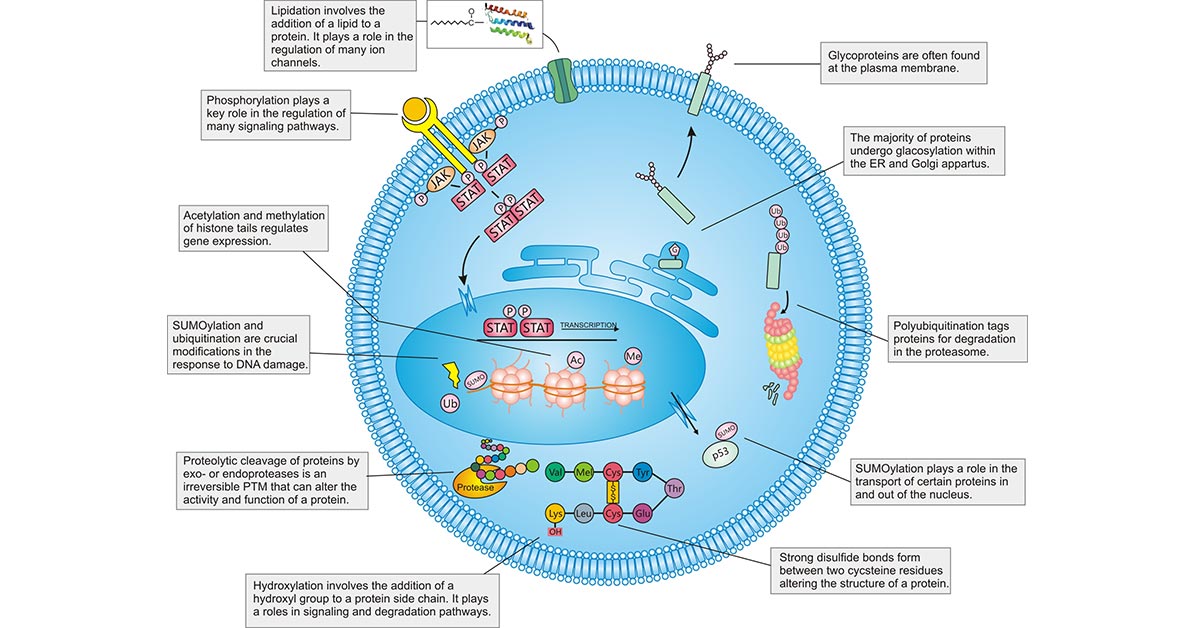
Figure 1: PTMs within the mammalian cell.
The picture is cited from: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8072572/
Table 1: Mechanisms and functions of different PTMs
| Types of PTMs | Mechanism | Targeted Sites | Function | |||||
|---|---|---|---|---|---|---|---|---|
| Phosphorylation | reversible | Transferring a phosphate group from adenosine triphosphate to the receptor residues by kinase enzymes. | Mainly happens on Ser, Thr, Tyr and His residues of the targeted protein, but also seen on Pro, Arg, Asp, and Cys residues. |
Involved in key cellular processes such as replication, transcription, environmental stress response, cell movement, cell metabolism, apoptosis, and immunological responsiveness. |
||||
| Acetylation | Nα-acetylation | irreversible | Acetyltransferases use acetyl CoA as a cofactor for adding an acetyl group (COCH3) to the ε-amino group of lysine side chains. | More reported on Lysine residue, but also occurs on Lys, Ala, Arg, Asp, Cys, Gly, Glu, Met, Pro, Ser, Thr and Val residues. | Plays an important role in biological processes such as chromatin stability, protein–protein interaction, cell cycle control, cell metabolism, nuclear transport and actin nucleation [5] [6]. | |||
| Nε-acetylation | reversible | |||||||
| O-acetylation | reversible | |||||||
| Ubiquitylation | reversible | A covalent bond befalls between the C-terminal of an active ubiquitin protein and Nε of a lysine residue of the protein. | Occur on all 20 amino acids, more frequently on lysine. | Mainly degrade intracellular proteins via the ubiquitin–proteasome pathway in all tissues. | ||||
| Methylation; | reversible | Addition of a methyl group, most often at lysine or arginine residues of the targeted protein. | Occurs on the Lys, Arg, Ala, Asn, Asp, Cys, Gly, Glu, Gln, His, Leu, Met, Phe and Pro residues in target proteins. | Most frequently seen in histone modification. Associated with fine tuning of various biological processes ranging from transcriptional regulation to epigenetic silencing via heterochromatin assembly. | ||||
| Glycosylation | N-glycosylation | reversible | N-linked glycans are attached to a nitrogen of asparagine or arginine side-chains. | Occurs more frequently on Ser, Thr, Asn and Trp residues in proteins and lipoproteins, but also occurs on Ala, Arg, Asp, Ile, Lys, Val, Glu, Pro, Tyr, Cys and Gly residues. | Participating in cell adhesion, cell–cell and cell–matrix interactions, molecular trafficking, receptor activation, protein solubility effects, protein folding and signal transduction, protein degradation, and protein intracellular trafficking and secretion [7-9]. | |||
| O-glycosylation | O-linked glycans attached to the hydroxyl oxygen of serine, threonine, tyrosine, hydroxylysine, or hydroxyproline side-chains, or to oxygens on lipids such as ceramide. | |||||||
| C-glycosylation | A sugar is added to a carbon on a tryptophan side-chain. | |||||||
| phosphoglycosylation | Phosphoglycans are linked through the phosphate of a phosphoserine. | |||||||
| glypiation | A GPI anchor is introduced and links proteins to lipids through glycan linkages. | |||||||
| SUMOylation | reversible | SUMO (SUMO1, SUMO2, and SUMO3) is connected to a lysine residue in substrate protein by covalent linkage via three enzymes, namely SAE1 (activating), UBC9 (conjugating) and ligase PIAS1. | lysine residue | Necessary to maintain genome integrity, transcriptional regulation, gene expression, and signal transduction in cells [10]. Regulating DNA damage repair, immune responses, carcinogenesis, cell cycle progression, and apoptosis. | ||||
| Lipidation | Palmitoylation | reversible | Covalent addition of palmitate to a cysteine via a thioester linkage. | Occurs on the Cys, Gly, Ser, Thr and Lys residues. | Plays critical roles in protein function regulation, protein–protein interaction, membrane–protein associations, neuronal development, signal transduction, apoptosis, and mitosis [11]. | |||
| Myristoylation | irreversible | Myristic acid is attached to the N-terminal glycine residue via a covalent bond. | Occurs more frequently on Gly and less frequently on Lys. | Regulating the protein structure maturation, signaling, extracellular communication, metabolism and the catalytic activity of the enzymes. | ||||
| Prenylation | Farnesylation | irreversible | Addition of farnesyl pyrophosphate to cysteine residues. | Occurs on cysteine and near the carboxyl-terminal end of the substrate protein. | Facilitating protein–protein interactions, endocytosis modulation, cell growth, differentiation, proliferation, and protein trafficking. | |||
| geranylation | irreversible | Addition of geranylgeranyl pyrophosphate to cysteine residues. | ||||||
| Sulfation | irreversible | TPST1 or TPST2 governs the transfer of an activated sulfate from 3-phospho adenosine 5-phosphosulfate to tyrosine residues within acidic motifs of polypeptides. | Occurs on Tyr, Cys, and Ser of the targeted proteins. | Has vital roles in protein–protein interactions, leukocyte rolling on endothelial cells, visual functions and viral entry into cells. | ||||
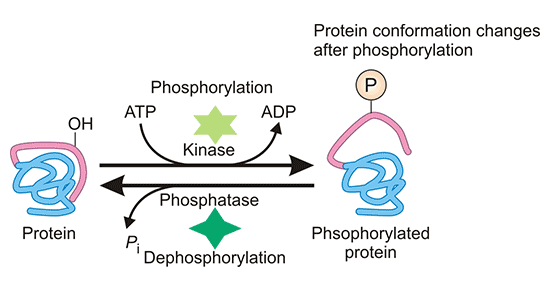
Figure 2: Phosphorylation of proteins
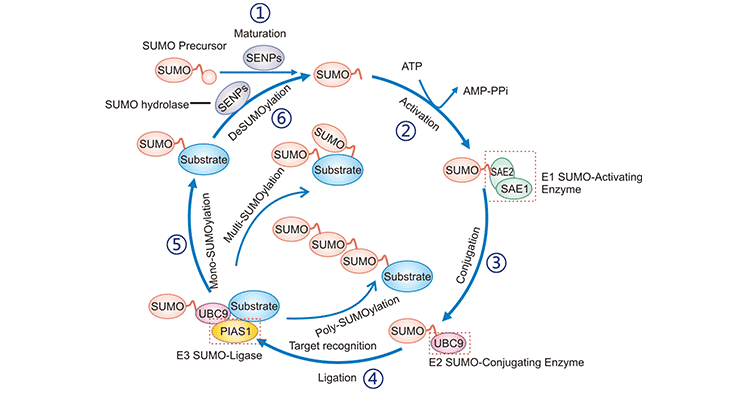
Figure 3: SUMOylation catalytic cycle
The picture is cited from: https://www.caister.com/cimb/v/v40/189.pdf
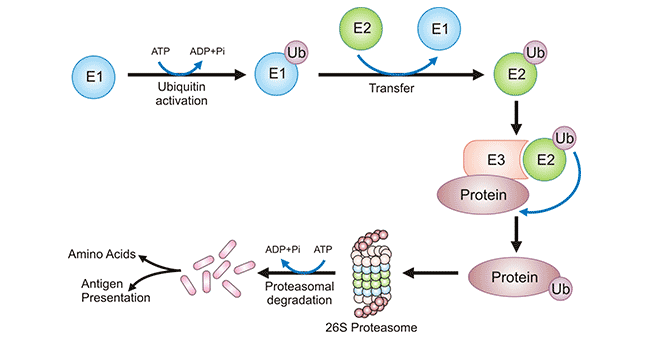
Figure 4: Ubiquitylation process of a protein and its degradation
The picture is sourced from: https://www.frontiersin.org/articles/10.3389/fmicb.2018.02738/full
PTMS of proteins plays a significantly important role in living organisms, which complicate the protein structure, perfect protein functions, refine the regulation, and makes the protein role more specific. PTMs expand the variability and diversity of protein structures and introduce new functionalities by widening the range of structures and physico-chemical properties of modified amino acid residues [12]. Since distinct PTMs can cause different alterations in physico-chemical properties of a protein [13], different modifications can confer the same protein distinct functionalities. And a protein can have different kinds and quantities of PTMs.
PTMs affect protein functions in various aspects. They may impact enzyme function and assembly [14], protein activity, protein longevity, protein–protein interactions [15], cell–cell/matrix interactions, molecular trafficking, receptor activation, protein solubility [16-18], protein folding [19], protein location, protein stability, and protein electrophilicity, among other protein behaviors and characteristics. As a result, these modifications play a role in a variety of biological processes, including signal transduction [20] [21], gene expression modulation, DNA repair, cell survival, and cell cycle control.
Detecting post-translational modifications, learning about how they function, affect the proteome, and govern the genome will vastly advance our understanding of genetics and epigenetics. PTM identification is important to elucidate the mechanism of complex cellular processes and diseases. Many experimental methods have been used to detect PTMs, including western blotting (WB), reverse phase protein arrays (RPA), immunoprecipitation (IP)-based Method, mass spectrometry (MS), in vitro assays, and immunofluorescence. They differ widely in terms of ease of use, sophistication, cost, and results delivery. Two or more methods may be combined to identify, validate, and mechanistic characterize a PTM for a protein of interest.
Table 2: Differences between different detection methods of PTMs
| Methods | Mechanism | Advantages | Disadvantages |
|---|---|---|---|
| Western blotting (WB) | Different proteins are separated from complex cellular lysates according to the molecular weight within an acrylamide gel and then the posterior is transferred into a membrane for blotting with primary and secondary antibodies for the selected PTMs. | May be able to investigate endogenous alterations of PTMs. No specific tools are required. | Require specific antibodies against the targeted protein. No site specificity. False negatives are possible due to PTM modifications may block the antibody binding site of the target protein. |
| Reverse phase protein arrays (RPA) | Use a platform that immobilizes cell lysates in small spots and allows antibodies that specifically recognize the PTM of interest to quantify PTMs of preselected proteins. | A sensitive antibody based proteomic approach that enables simultaneous quantification of multiple proteins and post-translational modifications across a multiple sample set. Relative high reproducibility, sensitivity, and robustness. | Antibodies need to be screened in advance and validated their specificity. The ability to collect samples and print viscous samples is a limitation of RPA technology. |
| Immunoprecipitation (IP)-based Method | Antibodies immobilized on a solid support matrix (e.g. agarose resin) bind to the protein of interest, and other proteins in the complex lysate are washed out. Use elution buffers to wash off the captured proteins and isolate them in a concentrated volume. The separated proteins are further analyzed by WB or MS. | Suitable for the enrich for specific, low-abundance PTMs on a target protein of interest. Optimized, high-quality IP reagents increase the accuracy of results. | Further analyse need other approaches like WB or MS. |
| Mass spectrometry (MS) | Digest the protein lysate of interest by a specific protease, usually trypsin, enrich the specific PTM, and analyze by LC-MS/MS. After collecting the data, computational algorithms are employed to identify peptides and proteins, as well as their relative quantification. Used to detect PTMs substrates and to map PTMs sites. |
Can detect large numbers of modified proteins with a high level of sensitivity and specificity. |
Time-consuming and highly PTM specific. Requires mass spectrometer and related experts who can familiarly operate the instrument. Limitations and deviations of mass spectrometers. |
| Immunofluorescence; | May be useful to investigate global and spatial changes in a PTM profile in tissues or cells. May be a useful biological readout. | Can be used to examine the localization of proteins throughout the cell and to determine localization of specific forms of proteins in response to different cell conditions. | Not available to identify a target specific PTM. |
PTMs influence almost all aspects of normal cell biology and fine-tune various molecular functions. Misregulation of PTMs can lead to the dysfunction of important biological processes including cell growth, metabolism, differentiation, and apoptosis, which contributes to the development and progression of diseases. Therefore, identifying and understanding PTMs is critical in the study of cell biology and disease treatment and prevention.
The disturbance in the pathway of phosphorylation has been linked to cancer, heart disease, Alzheimer's disease, and Parkinson's disease, among other disorders [22]. SUMOylation influences cancer cell signaling and gene networks that modulate inflammation, immunity, and DNA damage, establishing a relationship between carcinogenesis, proliferation, metastasis, and apoptosis. Dysregulation of acetylated lysine can lead to severe diseases such as cancer, aging, immunological problems, neurological diseases, and cardiovascular diseases [5] [23] [24]. Dysfunction in the ubiquitin pathway can lead to diverse diseases such as different cancers, metabolic syndromes, inflammatory disorders, type 2 diabetes, and neurodegenerative diseases [25] [26]. Defects in methylation can cause numerous diseases such as cancer, mental retardation (Angelman syndrome), diabetes mellitus, lipofuscinosis, and occlusive disease [27] [28]. Glycosylation defect plays a significant role in the development of various diseases like cancer, liver cirrhosis, diabetes, HIV infection, Alzheimer's disease, and atherosclerosis [9] [29]. Palmitoylation dysfunction has been associated with many diseases including neurological diseases such as Huntington's disease, schizophrenia and Alzheimer's disease, and different cancers [30-32]. Disruption in prenylation contributes to the pathogenesis of cancer, cardiovascular and cerebrovascular disorders, bone diseases, progeria, metabolic diseases, and neurodegenerative diseases.
References
[1] Human Genome Sequencing Consortium (2004) Finishing the euchromatic sequence of the human genome. Nature 431: 931–945.
[2] Jensen ON. Modification-specific proteomics: characterization of post-translational modifications by mass spectrometry. Curr Opin Chem Biol 2004, 8: 33–41.
[3] Ramazi, S., Allahverdi, A. and Zahiri, J. Evaluation of post-translational modifications in histone proteins: a review on histone modification defects in developmental and neurological disorders [J]. J. Biosci. 2020, 45, 135.
[4] Blom, N., Sicheritz-Pontén, T., Gupta, R. et al. Prediction of post-translational glycosylation and phosphorylation of proteins from the amino acid sequence [J]. Proteomics, 2004, 4, 1633–1649.
[5] Choudhary, C., Kumar, C., Gnad, F. et al. (2009) Lysine acetylation targets protein complexes and co-regulates major cellular functions. Science, 325, 834–840.
[6] Wellen, KE., Hatzivassiliou, G., et al. ATP-citrate lyase links cellular metabolism to histone acetylation. Science., 2009, 324, 1076–80.
[7] Haltiwanger, R.S. and Lowe, J.B. Role of glycosylation in development. Annu. Rev. Biochem., 2004, 73, 491–537.
[8] Ohtsubo, K. and Marth, J.D. Glycosylation in cellular mechanisms of health and disease. Cell, 2006, 126, 855–867.
[9] Goulabchand, R., Vincent, T., Batteux, F. et al. Impact of autoantibody glycosylation in autoimmune diseases. Autoimmun. Rev., 2014, 13, 742–750.
[10] Han, Z. J., Feng, Y. H., et al. The post-translational modification, SUMOylation, and cancer (Review). Int. J. Oncol.52, 1081–1094 (2018).
[11] Aicart-Ramos, C., Valero, R.A. and Rodriguez-Crespo, I. Protein palmitoylation and subcellular trafficking. Biochim. Biophys. Acta (BBA) Biomembr., 2011, 1808, 2981–2994.
[12] Walsh, C. T., Garneau-Tsodikova, S., and Gatto, G. J. Jr. Protein posttranslational modifications: the chemistry of proteome diversifications. Angew. Chem. Int. Ed. Engl. 2005, 44, 7342–7372.
[13] Mann, M., and Jensen, O. N. Proteomic analysis of post-translational modifications. Nat. Biotechnol. 2003, 21, 255–261.
[14] Ryšlavá, H., Doubnerová, V., Kavan, D. et al. Effect of posttranslational modifications on enzyme function and assembly. J. Proteomics, 2013, 92, 80–109.
[15] Marshall, C. Protein prenylation: a mediator of protein-protein interactions. Science, 1993, 259, 1865–1867.
[16] Haltiwanger, R.S. and Lowe, J.B. Role of glycosylation in development. Annu. Rev. Biochem., 2004, 73, 491–537.
[17] Karve, T.M. and Cheema, A.K. Small changes huge impact: the role of protein posttranslational modifications in cellular homeostasis and disease. J. Amino Acids, 2011, 1–13.
[18] Ohtsubo, K. and Marth, J.D. Glycosylation in cellular mechanisms of health and disease. Cell, 2006, 126, 855–867.
[19] Del Monte, F. and Agnetti, G. Protein post-translational modifications and misfolding: new concepts in heart failure. Proteomics Clin. Appl., 2014, 8, 534–542.
[20] Deribe YL, Pawson T, Dikic I. Post-translational modifications in signal integration. Nat Struct Mol Biol 2010, 17: 666–672.
[21] Zhao S, Xu W, et al. Regulation of cellular metabolism by protein lysine acetylation. Science 2010, 327: 1000–1004.
[22] Nsiah-Sefaa, A. and McKenzie, M. Combined defects in oxidative phosphorylation and fatty acid β-oxidation in mitochondrial disease. Biosci. Rep., 2016, 36, e00313.
[23] Falkenberg, K.J. and Johnstone, R.W. Histone deacetylases and their inhibitors in cancer, neurological diseases and immune disorders. Nat. Rev. Drug Discov., 2014, 13, 673.
[24] Park, G., Tan, J., Garcia, G. et al. Regulation of histone acetylation by autophagy in Parkinson disease. J. Biol. Chem., 2016, 291, 3531–3540.
[25] Micel, L.N., Tentler, J.J., Smith, P.G. et al. Role of ubiquitin ligases and the proteasome in oncogenesis: novel targets for anticancer therapies. J. Clin. Oncol., 2013, 31, 1231.
[26] Popovic, D., Vucic, D. and Dikic, I. Ubiquitination in disease pathogenesis and treatment. Nat. Med., 2014, 20, 1242–1253.
[27] Robertson, K.D. DNA methylation and human disease. Nat. Rev. Genet., 2005, 6, 597.
[28] Sun, G.-D., Cui, W.-P., Guo, Q.-Y. et al. Histone lysine methylation in diabetic nephropathy. J. Diabetes Res., 2014, 1–9.
[29] Lauc, G., Huffman, J.E., Pučić, M. et al. Loci associated with N-glycosylation of human immunoglobulin G show pleiotropy with autoimmune diseases and haematological cancers. PLoS Genet., 2013, 9, e1003225.
[30] Li, S., Li, J., Ning, L. et al. In silico identification of protein S-palmitoylation sites and their involvement in human inherited disease. J. Chem. Inf. Model,2015, 55, 2015–2025.
[31] Meckler, X., Roseman, J., Das, P. et al. Reduced Alzheimer's disease β-amyloid deposition in transgenic mice expressing S-palmitoylation-deficient APH1aL and nicastrin. J. Neurosci., 2010, 30, 16160–16169.
[32] Resh, M.D. Palmitoylation of proteins in cancer. Biochem. Soc. Trans., 2017, 45, 409–416.


Comments
Leave a Comment