Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
Myosin-regulated light chains (MLCs/MRLCs) constitute crucial elements within Myosin II, exerting significant influence on diverse life processes such as muscle contraction and cell division. Recent investigations have unveiled multiple isoforms of MLCs, including MYL9, MYL12A and MYL12B, sharing high sequence homology. Specifically, MYL12A and MYL12B form complexes with actin heavy chains, vital for regulating cell movement. Their phosphorylation status adjusts the actin complex activity, impacting muscle contraction strength and cell movement speed. Recent studies emphasize MYL12A and MYL12B's key roles in various diseases, linking their phosphorylation levels to conditions such as myocardial injury, hypertrophy, and cancer. Thus, MYL12A and MYL12B emerge as promising markers and potential targets in various diseases.
1. What is Myosin Regulatory Light Chain (MRLC)?
2. What are MYL12A and MYL12B?
3. What is the Ligand Associated with MYL12A and MYL12B?
6. Prospects for Clinical Research of MYL12A and MYL12B
7. CUSABIO MYL12A and MYL12B Recombinant Proteins & Antibodies for Research Use
Myosins have various components, including Myosin Heavy Chains and Myosin Light Chains (Click to see What is the Myosin Family?). The Myosin Light Chains are divided into basic MLC (MLC I) and regulatory MLC (MLC II or Myosin Regulatory Light Chain, MRLC). The Myosin Heavy Chain (MHC) acts as the motor domain, with MLC I stabilizing its structure, while MLC II primarily regulates myosin activity. MRLC can be classified into two major types: class I MRLC found in muscles (MLC1, MLC2, MLC3, MYL4, MYL5, MYL6, MYL7, etc), and class II MRLCs present in non-muscle cells (such as MYL9, MYL12A, MYL12B). Although structurally similar, class I and class II MRLCs differ in phosphorylation sites, with the former phosphorylated mainly at Ser19 and the latter at Ser17 [1-4].
MYL12A (MLCB, MRLC3, RLC) and MYL12B (MRLC2, MYLC2B, MLC2, MLC-2A, MLC-2) are regulatory components in the myosin light chain family (MLC II). They control myosin activity by phosphorylating the head region of myosin heavy chains. Both classes share an N-terminal motor domain, a "neck" lever arm, and a C-terminal tail. The N-terminal motor domain binds to actin, forming specific structures. The lever arm, with one to six isoleucine-glutamine (IQ) motifs, enhances function by interacting with calmodulin, causing changes in the motor domain. Conversely, the diverse C-terminal tail is crucial for protein interactions, serving as a key regulatory hotspot (Figure 1) [5-7].
Despite their similar amino acid sequences, MYL12A and MYL12B are two regulatory light chains with slight differences in expression and function. MYL12A is primarily expressed in smooth muscle, while MYL12B is widely distributed in various tissues, including skeletal muscle [7-11]. Additionally, they vary in ATPase activity, with MYL12A exhibiting lower activity, resulting in slower muscle contraction, and MYL12B displaying higher activity, leading to faster muscle contraction [8-12]. Notably, MYL12B serves as a substrate for serine/threonine kinase in the MAPK pathway, playing a pivotal role in muscle contraction and actin fibrosis in non-muscle cells [12-15].
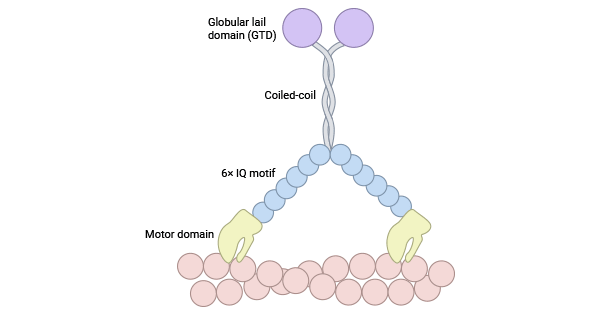
Figure 1. Myosin II structure [7]
Recent studies show that myosin light chains, including MYL12 (MYL12A, MYL12B) and MYL9, are new ligands for CD69. Blocking CD69-MYL9/12 interaction reduces allergic airway inflammation in mouse asthma models. In airway inflammation, MYL9/12 proteins are mainly found on the vascular surface. Inhibiting CD69-MYL9/12 interaction decreases leukocyte infiltration and improves airway inflammation, highlighting the role of MYL9/12 in recruiting CD69-expressing leukocytes to inflamed tissues. Analysis of nasal polyps in patients with eosinophilic chronic rhinosinusitis shows increased MYL9/12 expression in inflammatory lesions. This study identifies MYL12A, MYL12B, and MYL9 as novel ligands for CD69, playing a crucial role in recruiting activated CD69+ T cells to inflamed tissues, suggesting a potential therapeutic target for persistent inflammatory airway diseases [16-18].
Myosin function centers around the phosphorylation and dephosphorylation of the myosin light chain, regulated by two kinase systems: myosin light chain kinase (MLCK) and Rho-kinase (ROCK). Despite multiple signaling pathways influencing cytoskeletal remodeling and cell movement through MYL12A and MYL12B, our understanding is limited and requires further exploration. This brief summary highlights two key systems: MLCK and ROCK [19-23].
MLCK is a crucial regulator of myosin light chain phosphorylation, consisting of four kinases, including MLCK1/smMLCK and MLCK2. It operates as a calcium-dependent protein kinase, activated by increased intracellular Ca2+ levels binding to calmodulin. This activation leads to the phosphorylation of Ser19 and Thr18 on MLC II, activating the ATPase in myosin's head. This process triggers actin-myosin interactions, regulating various biological functions involving myosin. MLCK can also phosphorylate myosin light chains without calcium dependency, primarily through autophosphorylation at high concentrations. However, the main regulatory pathway for myosin light chain phosphorylation is through calcium-dependent MLCK (Figure 2) [22].
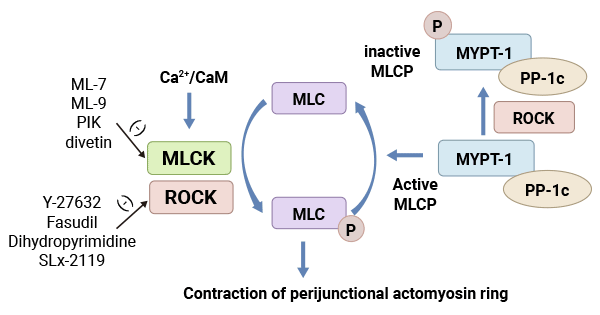
Figure 2. Myosin light chain kinase system (MLCK) [22]
ROCK, a serine/threonine protein kinase, has two isoforms—ROCKI and ROCKII—in mammalian cells. Rho, a member of the Ras superfamily with GTPase activity, serves as the upstream activator of Rho kinase. When activated by Rho, ROCK undergoes phosphorylation at multiple amino acid sites, initiating downstream phosphorylation/dephosphorylation reactions. Myosin phosphatase, activated by Rho/Rho kinase signals, phosphorylates its regulatory subunit, leading to its inactivation. This inactivated state prevents myosin phosphatase from dephosphorylating myosin light chain (MLC), causing an increase in intracellular cytoplasmic MLC phosphorylation levels and promoting actin-myosin interactions (Figure 3) [23].
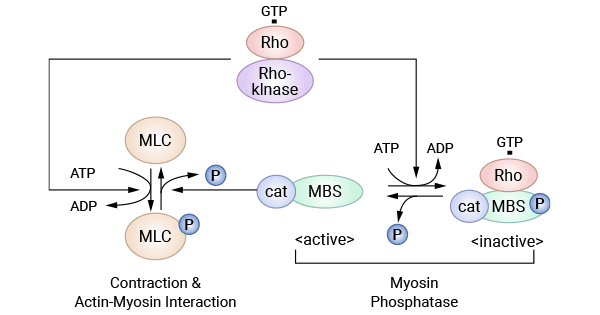
Figure 3. Rho kinase system [23]
Myosins, integral to cellular functions, regulate their activity through the phosphorylation and dephosphorylation of light chains, playing a crucial role in various diseases. Limited research exists on the involvement of MYL12A and MYL12B in diseases, but some studies indicate associations with cardiac diseases, cancer, intestinal barrier function, ischemic encephalopathy, and other health conditions.
Reduced phosphorylation of MRLC (MYL12A and MYL12B) is associated with cardiac torsion, myocardial dysfunction, and symptoms of dilated cardiomyopathy. In hypertrophic cardiomyopathy, MYLK3 knockout or mutation is linked to myocardial hypertrophy, and decreased MRLC phosphorylation may contribute to its development. Studies on heart failure indicate that MYLK3 knockout mice show cardiac function issues, and heart failure patients exhibit a significant decrease in MYL2 phosphorylation levels. These findings highlight the vital role of MRLC phosphorylation in the development and progression of heart disease [24-28].
Previous studies indicate that bacterial infiltration resulting from a disrupted intestinal barrier triggers immune responses, activating receptors like TLR4, TLR, and NLRP3. This process leads to cytokine release, including IL-13 and IFN-γ. Excessive IFN-γ has been associated with increased permeability, involving the MLCK pathway. However, the specific role of the MLCK/p-MLC2 pathway in intestinal barrier dysfunction during acute pancreatitis (AP) remains unclear. A study showed a significant increase in MLCK and p-MLC2 protein levels in ileal tissues during AP, particularly within the first 24 hours. This suggests that in acute pancreatitis, the activation of MLCK, phosphorylation of MLC2, and involvement of the MLCK/p-MLC2 pathway may contribute to intestinal barrier dysfunction [35-36].
Research indicates that the myosin light chain serves as a crucial tumor marker, influencing tumor proliferation, invasion, and migration through the regulation of myosin activity via phosphorylation and dephosphorylation [29]. MYPT1, the regulatory subunit of myosin phosphatase, catalyzes myosin light chain dephosphorylation, participating in the regulation of cellular motility, including processes like smooth muscle contraction. MYPT1 also inhibits the proliferation and metastatic activity of gastric cancer cells by affecting the phosphorylation of RhoA [30]. In highly malignant invasive ductal carcinomas of the breast, both MYL12A and MYL12B, isoforms of myosin light chain, are strongly and positively expressed, suggesting a close association with breast cancer development. In breast cancer tissues with high metastatic potential, the inhibition of MYPT1 expression increases MRLC phosphorylation, enhancing tumor cell motility and promoting metastasis [4, 31-34].
Myosin regulatory light chains (MYL12A and MYL12B) are linked to various diseases. In cerebral ischemia-reperfusion, using a CCR5 antagonist (DAPTA) reduced expression of ROCK2 and P-MLC2 (Ser19) proteins in rat brains, leading to milder neurological deficits and cerebral infarction compared to untreated rats, suggesting CCR5's impact on MLC phosphorylation via the ROCK pathway [37]. In Fcγ receptor-mediated phagocytosis, MYL9 is the primary MRLC isoform controlling uptake, while actin light chain 12A or 12B is not involved [38].
TRE17 oncoproteins directly bind MLC2 proteins and indirectly interact with the Rho protein family (Cdc42 and Rac1), regulating cytoskeletal actin remodeling [39]. Additionally, smooth muscle myosin light chain kinase (sm MLCK)-mediated phosphorylation of MYL12A/MYL12B induces cellular morphology changes in cochlear hair cells (HCs), potentially impacting auditory function [40].
The myosin family, particularly myosin-regulated light chain RLCs (MYL12A and MYL12B), has garnered attention in disease research. These components play key roles in pathways like MLCK and Rho kinase, influencing processes such as cell proliferation, metastasis, and inflammation. Additionally, they act as ligands for CD69, impacting immune responses. Phosphorylation of myosin regulatory light chains (MRLCs) within the myosin family has vital biological functions in various muscle tissues, suggesting significant implications for diverse diseases. Ongoing research on MYL12A and MYL12B roles in myocardial injury, tumors, autoimmune diseases offers new perspectives for clinical studies, making them crucial targets for future research.
MYL12A and MYL12B, crucial components of the myosin family, play key roles in biological processes like cell proliferation, metastasis, and inflammation through pathways like MLCK and Rho kinase. They also serve as ligands for CD69, impacting immune responses. The phosphorylation of myosin regulatory light chains (RLCs) within the myosin family has significant implications for various diseases. Recent research underscores their involvement in conditions like myocardial injury, tumors, and autoimmune diseases, making MYL12A and MYL12B promising targets for clinical research.
To fully support researchers and pharmaceutical companies in their research on MYL12A and MYL12B in hearing, heart disease, intestinal barrier function, ischemic encephalopathy, cancer and other diseases, CUSABIO presents MYL12A and MYL12B active proteins to support your research on the mechanism of MYL12A and MYL12B or their potential clinical value.
CUSABIO MYL12A and MYL12B proteins
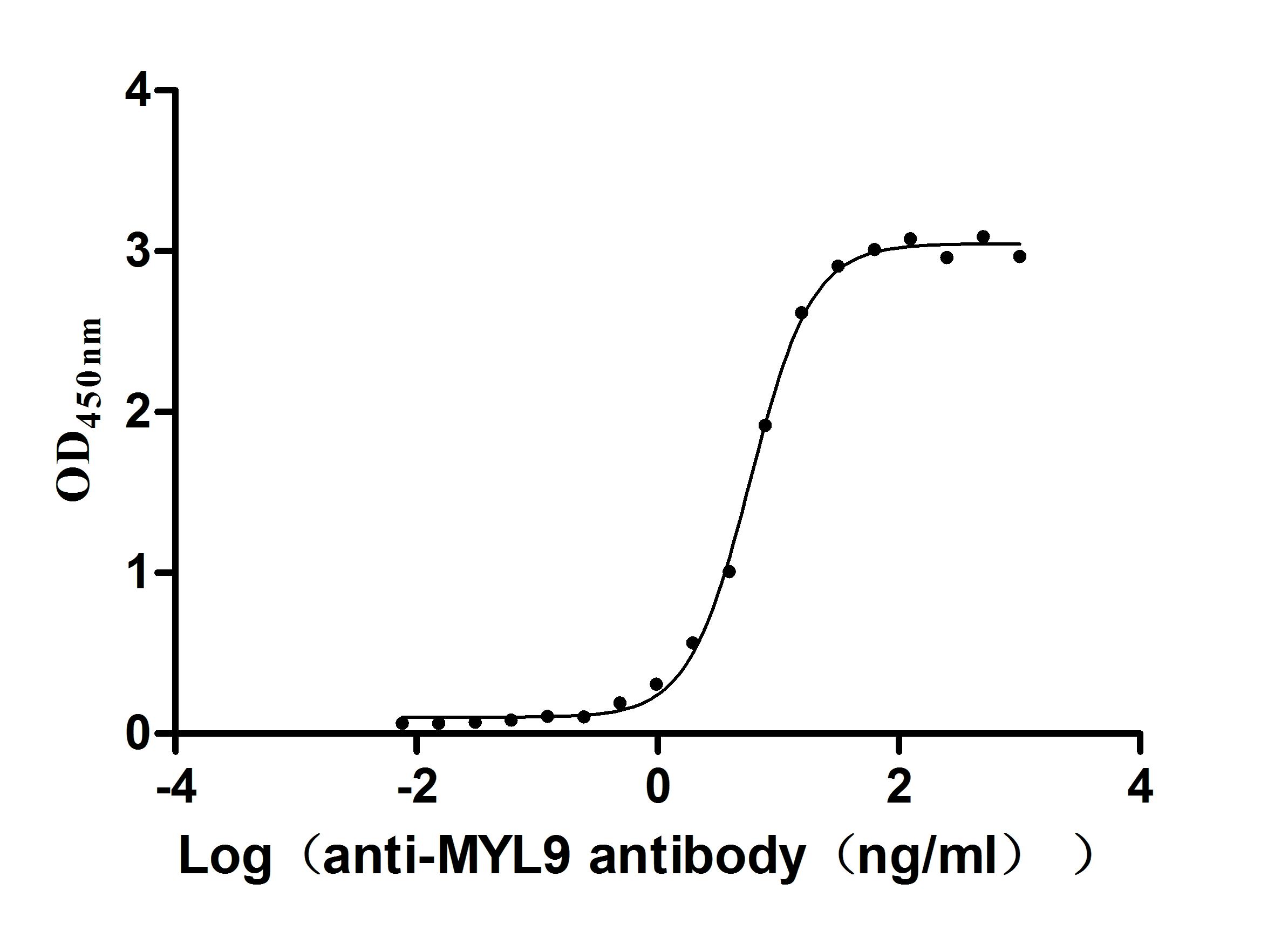
● High Purity: ≥ 95% as determined by SDS-PAGE.
● Verified Activity by a functional ELISA: immobilized Human MYL12A at 2μg/mL can bind Anti-MYL9 recombinant antibody (CSB-RA015318MA1HU). The EC50 is 5.325-6.456 ng/mL.
● High Purity: ≥ 95% as determined by SDS-PAGE.
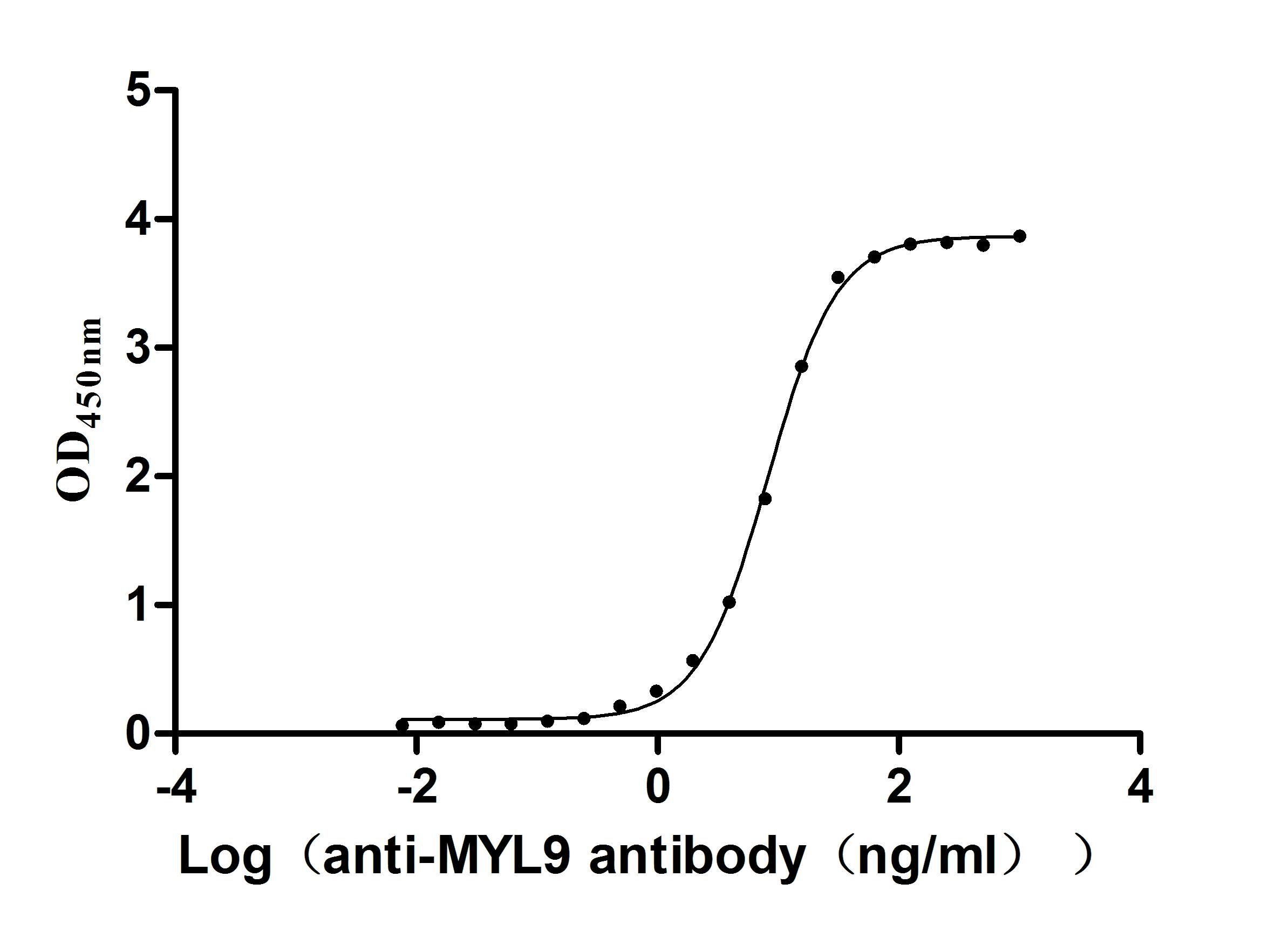
● Verified Activity by a functional ELISA: Immobilized Human MYL12B at 2μg/mL can bind Anti-MYL9 recombinant antibody (CSB-RA015318MA1HU). The EC50 is 7.760-8.646 ng/mL.
References
[1] Cao, Lichuang, et al. "Phosphorylation of myosin regulatory light chain affects actomyosin dissociation and myosin degradation." International Journal of Food Science & Technology 54.6 (2019): 2246-2255.
[2] Gao, Xing, et al. "Dephosphorylation of myosin regulatory light chain modulates actin–myosin interaction adverse to meat tenderness." International Journal of Food Science & Technology 52.6 (2017): 1400-1407.
[3] Cao, Lichuang, et al. "Phosphorylation of myosin regulatory light chain at Ser17 regulates actomyosin dissociation." Food Chemistry 356 (2021): 129655.
[4] Aguilar-Cuenca, Rocío, et al. "Tyrosine phosphorylation of the myosin regulatory light chain controls non-muscle myosin II assembly and function in migrating cells." Current Biology 30.13 (2020): 2446-2458.
[5] Sweeney, H. Lee, and Erika LF Holzbaur. "Motor proteins." Cold Spring Harbor Perspectives in Biology 10.5 (2018): a021931.
[6] Mackay, Charles Edward. SrcFK is a key mediator of oxidant signalling pathways in Pulmonary Vascular Smooth Muscle. Diss. King's College London, 2016.
[7] Haraguchi, Takeshi, et al. "Discovery of ultrafast myosin, its amino acid sequence, and structural features." Proceedings of the National Academy of Sciences 119.8 (2022): e2120962119.
[8] Kumar, C. Chandra, et al. "Characterization and differential expression of human vascular smooth muscle myosin light chain 2 isoform in nonmuscle cells." Biochemistry 28.9 (1989): 4027-4035.
[9] Wang, Shaoxun, et al. "Down‐Regulation of Gamma‐Adducin Disrupts the Actin Cytoskeleton in FHH rats and May Contribute to the Development of Hypertension‐induced Renal Injury." The FASEB Journal 32 (2018): 721-10.
[10] Dabrowska, Magdalena, Marek Skoneczny, and Wojciech Rode. "Functional gene expression profile underlying methotrexate-induced senescence in human colon cancer cells." Tumor Biology 32 (2011): 965-976.
[11] Jiang, Yuhui, et al. "PKM2 phosphorylates MLC2 and regulates cytokinesis of tumour cells." Nature communications 5.1 (2014): 5566.
[12] Iwasaki, Takahiro, et al. "Diphosphorylated MRLC is required for organization of stress fibers in interphase cells and the contractile ring in dividing cells." Cell structure and function 26.6 (2001): 677-683.
[13] Gutjahr, Marc C., Jérémie Rossy, and Verena Niggli. "Role of Rho, Rac, and Rho-kinase in phosphorylation of myosin light chain, development of polarity, and spontaneous migration of Walker 256 carcinosarcoma cells." Experimental cell research 308.2 (2005): 422-438.
[14] Parker, Robert, et al. "Phosphoproteomic analysis of cell-based resistance to BRAF inhibitor therapy in melanoma." Frontiers in oncology 5 (2015): 95.
[15] Wang, Shibo, et al. "Myosin light chain kinase mediates intestinal barrier dysfunction following simulated microgravity based on proteomic strategy." Journal of proteomics 231 (2021): 104001.
[16] Hayashizaki, Koji, et al. "Myosin light chains 9 and 12 are functional ligands for CD69 that regulate airway inflammation." Science immunology 1.3 (2016): eaaf9154-eaaf9154.
[17] Kimura, Motoko Y., et al. "A new therapeutic target: the CD69-Myl9 system in immune responses." Seminars in immunopathology. Vol. 41. Springer Berlin Heidelberg, 2019.
[18] Yokoyama, Masaya, et al. "Myosin light chain 9/12 regulates the pathogenesis of inflammatory bowel disease." Frontiers in Immunology 11 (2021): 594297.
[19] Sun, Jie, et al. "Distinct roles of smooth muscle and non-muscle myosin light chain-mediated smooth muscle contraction." Frontiers in Physiology 11 (2020): 593966.
[20] Isobe, Kiyoshi, et al. "CRISPR-Cas9/phosphoproteomics identifies multiple noncanonical targets of myosin light chain kinase." American Journal of Physiology-Renal Physiology 318.3 (2020): F600-F616.
[21] Orgaz, Jose L., et al. "Myosin II reactivation and cytoskeletal remodeling as a hallmark and a vulnerability in melanoma therapy resistance." Cancer Cell 37.1 (2020): 85-103.
[22] Jin, Younggeon, and Anthony T. Blikslager. "The regulation of intestinal mucosal barrier by myosin light chain kinase/rho kinases." International Journal of Molecular Sciences 21.10 (2020): 3550.
[23] Kaibuchi, Kozo, Shinya Kuroda, and Mutsuki Amano. "Regulation of the cytoskeleton and cell adhesion by the Rho family GTPases in mammalian cells." Annual review of biochemistry 68.1 (1999): 459-486.
[24] Scruggs, Sarah B., and R. John Solaro. "The significance of regulatory light chain phosphorylation in cardiac physiology." Archives of biochemistry and biophysics 510.2 (2011): 129-134.
[25] Ito, Masaaki, et al. "Regulation of myosin light-chain phosphorylation and its roles in cardiovascular physiology and pathophysiology." Hypertension Research 45.1 (2022): 40-52.
[26] Ding, Peiguo, et al. "Cardiac myosin light chain kinase is necessary for myosin regulatory light chain phosphorylation and cardiac performance in vivo." Journal of Biological Chemistry 285.52 (2010): 40819-40829.
[27] Huang, Jian, et al. "Myosin regulatory light chain phosphorylation attenuates cardiac hypertrophy." Journal of Biological Chemistry 283.28 (2008): 19748-19756.
[28] Davis, Julien S., et al. "The overall pattern of cardiac contraction depends on a spatial gradient of myosin regulatory light chain phosphorylation." Cell 107.5 (2001): 631-641.
[29] Tohtong, R., et al. "Dependence of metastatic cancer cell invasion on MLCK-catalyzed phosphorylation of myosin regulatory light chain." Prostate cancer and prostatic diseases 6.3 (2003): 212-216.
[30] Wu, Qian, et al. "Deficiency in myosin light-chain phosphorylation causes cytokinesis failure and multipolarity in cancer cells." Oncogene 29.29 (2010): 4183-4193.
[31] https://www.proteinatlas.org/ENSG00000101608-MYL12A/pathology
[32] Xiao, Xiao, et al. "Transformer with convolution and graph-node co-embedding: an accurate and interpretable vision backbone for predicting gene expressions from local histopathological image." Medical Image Analysis 91 (2024): 103040.
[33] Li, Yin-Chao, et al. "Oridonin suppress cell migration via regulation of nonmuscle myosin IIA." Cytotechnology 68 (2016): 389-397.
[34] Hosono, Yasuyuki, et al. "MYBPH inhibits NM IIA assembly via direct interaction with NMHC IIA and reduces cell motility." Biochemical and Biophysical Research Communications 428.1 (2012): 173-178.
[35] Du, Liwen, et al. "Inhibition of the MLCK/MLC2 pathway protects against intestinal heat stroke-induced injury in rats." Journal of Thermal Biology 116 (2023): 103655.
[36] Rath, Nicola, and Michael F. Olson. "Regulation of pancreatic cancer aggressiveness by stromal stiffening." Nature medicine 22.5 (2016): 462-463.
[37] Li, Laisi, et al. "Effects of CC-chemokine receptor 5 on ROCK2 and P-MLC2 expression after focal cerebral ischaemia–reperfusion injury in rats." Brain Injury 30.4 (2016): 468-473.
[38] Bright, Michael D., and Gad Frankel. "PAK4 phosphorylates myosin regulatory light chain and contributes to Fcγ receptor-mediated phagocytosis." The International Journal of Biochemistry & Cell Biology 43.12 (2011): 1776-1781.
[39] Oliveira, Andre M., and Margaret M. Chou. "The TRE17/USP6 oncogene: a riddle wrapped in a mystery inside an enigma." Frontiers in Bioscience-Scholar 4.1 (2012): 321-334.
[40] Oya, Ryohei, et al. "Phosphorylation of MYL12 by Myosin Light Chain Kinase Regulates Cellular Shape Changes in Cochlear Hair Cells." Journal of the Association for Research in Otolaryngology 22 (2021): 425-441.


Comments
Leave a Comment