Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
The vascular endothelial growth factor receptor (VEGFR) family has long been a focus of therapeutic interest. Among them, FLT1 (VEGFR-1) stands out for its unique role as a "bidirectional regulator." FLT1 is far more than a simple signal receiver; it can transmit signals via its membrane-bound form to promote endothelial cell migration and vascular remodeling, while its soluble form, sFLT1, acts as a ligand "trap," providing negative feedback to inhibit excessive vascular proliferation, thereby maintaining a crucial dynamic balance between pro-angiogenic and anti-angiogenic signals.
Recent studies indicate that the disruption of this balance is directly involved in various pathological processes, including liver fibrosis, impaired lung injury repair, osteoarthritis, and tumor progression. As its novel roles in immune regulation and drug resistance continue to be uncovered, FLT1 has emerged as a highly promising therapeutic target, attracting widespread attention from basic research to drug development. This article will systematically review the biological functions, signaling networks, and latest research advances of FLT1 in related diseases.
FLT1 (VEGFR-1) is a member of the vascular endothelial growth factor receptor (VEGFR) family and belongs to the typical Type III receptor tyrosine kinases. Its structure comprises seven extracellular immunoglobulin-like domains, a single transmembrane region, and an intracellular tyrosine kinase domain [1]. Alternatively spliced soluble FLT1 (sFLT1) lacks the transmembrane and kinase domains but retains VEGF-binding capacity, functioning as an endogenous VEGF signaling antagonist [2]. Crystallographic studies reveal that FLT1 and VEGF-A form a 1:2 complex, where the second immunoglobulin-like domain (D2) determines ligand recognition, while D3 participates in receptor dimerization and activation [3]. FLT1 exhibits differential affinity for VEGF family ligands, binding VEGF-B and Placental Growth Factor (PlGF) with higher affinity than VEGFR-2.
During angiogenesis, FLT1 exerts regulatory functions through dual mechanisms: membrane-bound FLT1, upon activation, recruits Gab1, subsequently activating the PI3K-Akt-mTOR pathway to promote endothelial cell migration [4]; whereas sFLT1 maintains vascular homeostasis by neutralizing free VEGF and inhibiting excessive angiogenesis [1]. Studies show that FLT1 knockout mice are embryonically lethal due to abnormal vascular development, a phenotype distinct from VEGFR-2 deficiency [2]. Phosphorylation of tyrosine 1210 in the FLT1 kinase domain specifically activates the HIF-1α-VEGF positive feedback loop, amplifying pro-angiogenic signals under hypoxic conditions [4]. Furthermore, FLT1 regulates Rho GTPase activity via non-canonical pathways, influencing endothelial cytoskeletal reorganization and lumen formation [3].
FLT1 function relies on dynamic glycosylation modifications. N-linked glycosylation affects receptor membrane localization and controls signaling intensity by modulating ligand-binding affinity. Under pathological conditions, metalloproteinase ADAM17-mediated shedding of the FLT1 extracellular domain generates sFLT1 fragments, a process significantly induced by TNF-α and oxidative stress [1]. FLT1 exhibits angiogenesis-independent functions in specific tissues, such as activating the TrkB pathway to regulate synaptic plasticity in neurons and inhibiting MMP-13 expression to delay matrix degradation in chondrocytes [3][4]. This multifunctionality positions FLT1 as a critical node connecting the vascular system and the microenvironment.
The vascular endothelial growth factor (VEGF) family includes VEGF-A, VEGF-B, VEGF-C, VEGF-D, and Placental Growth Factor (PlGF), with their receptors being FLT1 (VEGFR-1), KDR (VEGFR-2), and FLT4 (VEGFR-3) [5]. FLT1 was the first identified VEGF receptor, showing higher binding specificity for VEGF-B and PlGF compared to VEGFR-2, but possesses weaker kinase activity, primarily serving regulatory functions [1].
FLT1 and VEGFR-2 complementarily regulate angiogenesis: VEGFR-2 mediates endothelial proliferation, migration, and increased permeability, whereas FLT1 maintains vascular homeostasis by competitively binding VEGF-A to inhibit excessive VEGFR-2 activation, or by activating the PLCγ-PKC-MAPK pathway [5]. FLT1 knockout mice die due to excessive vascular hyperplasia, while transgenic mice expressing only the extracellular truncated form of FLT1 survive normally, indicating that its negative regulatory role primarily relies on a ligand trapping mechanism [3]. The PlGF-FLT1 axis can activate the PI3K/Akt pathway under pathological conditions, promoting vascular remodeling and playing a key role in ischemic diseases and tumor angiogenesis [6].
FLT1 cooperates with VEGFR-3 to regulate blood-lymphatic vessel balance. VEGFR-3 responds to VEGF-C/D to promote lymphangiogenesis, while FLT1 enhances the sensitivity of lymphatic endothelial cells to low concentrations of VEGF-C through the formation of heterodimers [1]. In wound repair models, FLT1 deficiency can lead to approximately a 40% reduction in lymphatic vessel density [3]. sFLT1 acts as a natural antagonist by binding circulating VEGF-A and PlGF to regulate vascular permeability, and its abnormally elevated levels are associated with the pathogenesis of pre-eclampsia [5]. Proteomic studies show that sFLT1 affinity for VEGF-A165 is 5-fold higher than that of VEGFR-2, but its inhibitory effect on VEGF-A121 is limited; this isoform selectivity offers targeted strategies for treating vascular diseases [2].
FLT1 expression is regulated at multiple levels:
FLT1 binding to VEGF induces dimerization, autophosphorylation, and recruitment of adaptor proteins like Akt, forming a signaling complex [4].
VEGF stimulation exhibits spatiotemporal specificity, enhancing vascular permeability in the short term and promoting angiogenesis in the long term [5]. FLT1 forms heterodimers with VEGFR-2 to coordinately regulate PI3K-Akt and ERK1/2 branches; Akt phosphorylation levels positively correlate with microvessel density. TNF-α can inhibit HIF-1α via NF-κB, blocking the FLT1-VEGF positive feedback loop and leading to impaired angiogenesis [8].
FLT1 forms complexes with VEGFR-2, with FLT1 biased towards the PLCγ-PKC-MAPK pathway and VEGFR-2 towards the PI3K-Akt pathway [4]. Under hypoxia, FLT1 competitively binds VEGF-B/PlGF, reducing excessive VEGFR-2 activation and maintaining vascular homeostasis [5]. The receptor ratio and phosphorylation status dynamically balance signal output, while changes in sFLT1 levels further fine-tune angiogenesis [2][5]. In cartilage repair, changes in the FLT1/VEGFR-2 ratio affect vascular invasion and MMP-9 secretion, providing a favorable microenvironment for tissue engineering [5].
Network pharmacology studies indicate that natural products and compound formulations can achieve vascular homeostasis by modulating FLT1 and its downstream signals [11].
FLT1-mediated angiogenesis promotes pathological vessel formation and extracellular matrix deposition in liver fibrosis [9]. In carbon tetrachloride-induced mouse models, FLT1 expression positively correlates with the degree of fibrosis, and inhibiting FLT1 reduces collagen deposition and hepatic stellate cell activation. A PDGF/VEGF autocrine loop accelerates the transformation of myofibroblasts, and the dual-target inhibitor RC28-E shows potential therapeutic value by reducing collagen deposition [12]. Single-cell sequencing reveals a high overlap between FLT1+ endothelial subpopulations and the expression of inflammatory factors IL-6 and TGF-β1 in fibrotic livers [13]. A lipotoxic environment leads to vascular dysfunction via upregulation of sFLT1 [14].
FLT1 plays a dual role in lung injury repair: maintaining vascular integrity in the acute phase, while excessive activation in the chronic phase leads to vascular abnormalities. Bleomycin-induced models show that FLT1 upregulates VEGF-A, promoting pathological capillary proliferation; RC28-E inhibits TGF-β1/Smad3 and improves lung function [12]. MSC-derived extracellular vesicles (MSC-EVs) carrying miR-214-3p inhibit FLT1 translation and promote alveolar epithelial regeneration [15][16]. FLT1 inhibitors show optimal efficacy when administered 7-14 days post-injury, balancing pro-angiogenic and anti-fibrotic needs.
FLT1 regulates subchondral vascularization, promoting endothelial migration and new blood vessel formation via HIF-1α signaling [3]. Moderate vascularization facilitates stem cell migration and nutrient supply, while sFLT1 competes with VEGF to inhibit excessive vascularization, maintaining homeostasis at the cartilage-bone interface [15]. FLT1 inhibition can reduce MMP-2/9 release, lowering the risk of matrix degradation [10]. Combining anti-FLT1 antibodies with hydrogel scaffolds enables structural cartilage repair [15].
In recent years, significant progress has been made in the development of FLT1-targeted drugs, particularly showing potential in anti-tumor immunity and reversing drug resistance. For example, studies have found that FLT1 activation can protect cancer cells from PARP inhibitor (PARPi) suppression via non-canonical pathways. Using FLT1 inhibitors (such as axitinib) or genetic knockout can inhibit the AKT signaling pathway, increase CD8+ T cell infiltration, and significantly reverse PARPi resistance. Currently, 10 small-molecule drugs are approved on the market. Selected other pipelines under investigation are listed in the table below:
| Drug | Mechanism of Action | Drug Type | Indications Under Investigation | R&D Institutions | Highest Phase |
|---|---|---|---|---|---|
| Lucitanib | FGFR1 antagonist | FGFR2 antagonist | VEGFR1 antagonist | VEGFR2 antagonist | VEGFR3 antagonist | Small Molecule | Colon Cancer | Nasopharyngeal Neoplasms | Nanjing Aidecheng Pharmaceutical Technology Co., Ltd. | Shanghai Haihe Pharmaceutical Co., Ltd. | Phase 3 |
| Cediranib | PDGFRβ inhibitor | VEGFR1 antagonist | VEGFR2 antagonist | VEGFR3 antagonist | c-Kit inhibitor | Small Molecule | Ovarian Endometrioid Adenocarcinoma | Fallopian Tube Clear Cell Adenocarcinoma | Ovarian Clear Cell Adenocarcinoma | Ovarian Mixed Epithelial Carcinoma, etc. | National Cancer Institute | AstraZeneca PLC | The University of Texas MD Anderson Cancer Center | Phase 3 |
| Zanzalintinib | AXL inhibitor | MerTK inhibitor | VEGFR1 antagonist | c-Met inhibitor | Small Molecule | Metastatic Clear Cell Renal Cell Carcinoma | Renal Cell Carcinoma | Metastatic Colorectal Cancer | Pancreatic Extra-Neuroendocrine Tumors, etc. | Exelixis, Inc. | Phase 3 |
| Sitravatinib | AXL inhibitor | RET inhibitor | Tie-2 antagonist | VEGFR1 antagonist | VEGFR2 antagonist | VEGFR3 antagonist | c-Met inhibitor | Small Molecule | Tumors | Advanced Squamous Non-Small Cell Lung Cancer | Metastatic Non-Squamous Non-Small Cell Lung Cancer | Liver Metastases, etc. | BeOne Medicines Ltd. | Mirati Therapeutics, Inc. | BeiGene (Guangzhou) Pharmaceutical Co., Ltd. | Phase 3 |
| VEGFR-1/2 peptide vaccine (Keio University) | VEGFR1 antagonist | VEGFR2 antagonist | Universal Antigen Vaccine | Synthetic Peptide Vaccine | Therapeutic Vaccine | Glioma | Neurofibromatosis | Keio University | Phase 2 |
| YP-01001 | FGFRs antagonist | RET inhibitor | VEGFR1 antagonist | VEGFR2 antagonist | VEGFR3 antagonist | Small Molecule | Solid Tumors | Advanced Malignant Solid Tumors | Thyroid Cancer | Chongqing Yaopharma Co., Ltd. | Shanghai Fosun Pharmaceutical (Group) Co., Ltd. | Phase 2 |
| CBP-4888 | VEGFR1 antagonist | siRNA | Preeclampsia | Comanche Biopharma Corp. | Phase 1 |
| Dual-targeting VEGFR1 and PD-L1 CAR-T cells (Sichuan University) | PDL1 modulator | VEGFR1 modulator | T-cell replacement | CAR-T | - | Sichuan University | Phase 1 |
| Hydroxychloroquine/Sorafenib | BRAF inhibitor | CRAF inhibitor | FLT3 inhibitor | PDGFRβ inhibitor | RET inhibitor | VEGFR1 antagonist | VEGFR2 antagonist | VEGFR3 antagonist | c-Kit inhibitor | Small Molecule | Ovarian Cancer | VG Life Sciences, Inc. | Phase 1 |
FLT1, as a key factor in angiogenesis and immune regulation, has become a research focus in various diseases. Its aberrant expression in CMS, PAH, OA, tumors, and other diseases suggests it is not only a potential diagnostic marker but also an important target for therapeutic intervention.
CUSABIO has developed a variety of FLT1-related research products, including recombinant proteins, antibodies, and ELISA kits, to assist your research into FLT1 mechanisms and drug development.
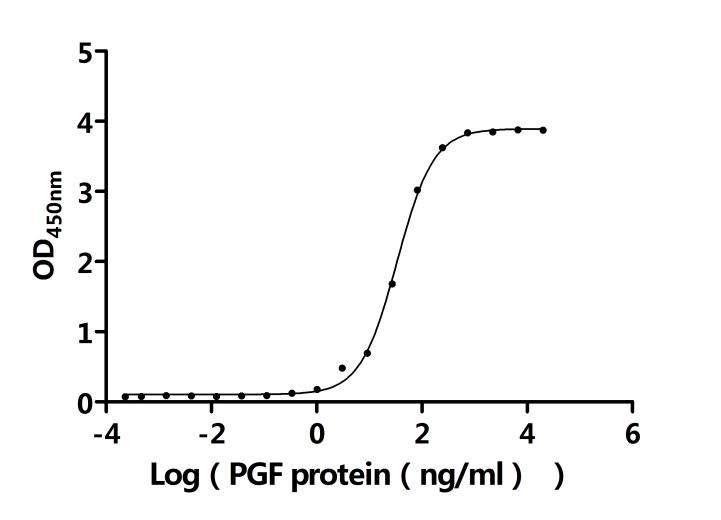
● FLT1 Recombinant Protein
● FLT1 Antibody
FLT1 Recombinant Monoclonal Antibody; CSB-RA940669A0HU

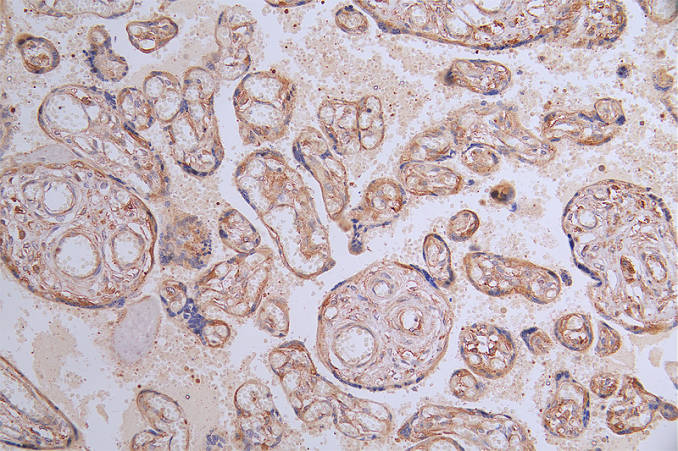
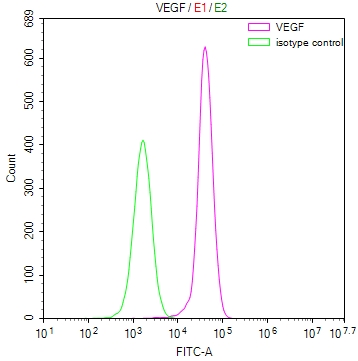
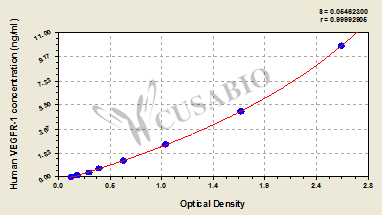
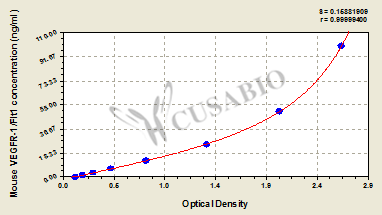
● FLT1 ELISA kit
References
[1] Yan‐Xia Zhang, Mi‐Rong Jing, Chunbo Cai, Shuai‐Gang Zhu, C. Zhang, Qi‐Meng Wang, Yuankun Zhai, Xin‐Ying Ji, Dongdong Wu. Role of hydrogen sulphide in physiological and pathological angiogenesis[J]. Cell Proliferation, 2022, 56(3).
[2] Fan-Wei Peng, Dake Liu, Qingwen Zhang, Yungen Xu, Lei Shi. VEGFR-2 inhibitors and the therapeutic applications thereof: a patent review (2012-2016)[J]. Expert Opinion on Therapeutic Patents, 2017, 27(9): 987-1004.
[3] Leopold F. Fröhlich. MicroRNAs at the Interface between Osteogenesis and Angiogenesis as Targets for Bone Regeneration[J]. Cells, 2019, 8(2): 121-121.
[4] Alexander Panossian, Ean‐Jeong Seo, Thomas Efferth. Novel molecular mechanisms for the adaptogenic effects of herbal extracts on isolated brain cells using systems biology[J]. Phytomedicine, 2018, 50: 257-284.
[5] Samanta Taurone, Filippo Galli, Alberto Signore, Enzo Agostinelli, Rudi Dierckx, Antonio Minni, Marcella Pucci, Marco Artico. VEGF in nuclear medicine: Clinical application in cancer and future perspectives (Review)[J]. International Journal of Oncology, 2016, 49(2): 437-447.
[6] Peiyu Liu, Dandan Sun, Shuchang Zhang, Shimei Chen, Xiaoqian Wang, Huiming Li, Wei Fang. PFKFB3 in neovascular eye disease: unraveling mechanisms and exploring therapeutic strategies[J]. Cell & Bioscience, 2024, 14(1).
[7] Maria Giovanna Scioli, Gabriele Storti, Federico D’Amico, Roger Rodríguez Guzmán, Federica Centofanti, Elena Doldo, Ela María Céspedes Miranda, Augusto Orlandi. Oxidative Stress and New Pathogenetic Mechanisms in Endothelial Dysfunction: Potential Diagnostic Biomarkers and Therapeutic Targets[J]. Journal of Clinical Medicine, 2020, 9(6): 1995-1995.
[8] Fengyan Jin, Xiangyu Zheng, Yanping Yang, Gang Yao, Long Ye, Thorsten R. Doeppner, Dirk M. Hermann, Haifeng Wang, Yun Dai. Impairment of hypoxia-induced angiogenesis by LDL involves a HIF-centered signaling network linking inflammatory TNFα and angiogenic VEGF[J]. Aging, 2019, 11(2): 328-349.
[9] Huajuan Jiang, Xulong Huang, Jiaxin Wang, Yongfeng Zhou, Chaoxiang Ren, Tao Zhou, Jin Pei. Hepatoprotective Effect of Medicine Food Homology Flower Saffron against CCl4-Induced Liver Fibrosis in Mice via the Akt/HIF-1α/VEGF Signaling Pathway[J]. Molecules, 2023, 28(21): 7238-7238.
[10] Weiwei Wu, Jiaying Zhao, Yingying Li, Wendi Sun, Xiaohong Ye, Xinyan Chang, Yao He, Qing Jin. Rg1 inhibits the VEGF-Flt1 signaling pathway and microvascular remodeling in knee osteoarthritis rats[J]. Phytotherapy Research, 2022, 36(11): 4991-5002.
[11] Limin Wang, Wenxia Zhang, Bing Zhao, Huijuan Li, Qing Liu, Ping Xu, Jianyong Lei, Zhibin Wang, Xinfeng Yan, Conghui Li. The effect of combined treatment with Salvia miltiorrhiza and Panax notoginseng on vascular regeneration and tissue repair of osteoarthritis[J]. Biomedicine & Pharmacotherapy, 2023, 164: 114869-114869.
[12] Siyuan Chen, Qiang Fu, Yinyan Tang, Ping Wang, Yali Zheng, Ling Ye, Feng Xu. Bispecific antibodies targeting VEGF/Ang-2 for the treatment of liver fibrosis[J]. Immunologic Research, 2023, 71(5): 682-693.
[13] Yuan Xu, Chen Wei, Shuang Liu, Han Sun, Chen Chen, Mengru Li, Zhuohang Li, Wenxing Hu, Xiaobing Wang, Xianmei Meng, Li Li, Hongbo Xin. Single-cell RNA sequencing uncovers heterogeneity and crosstalk of hepatic stellate cells in a CCl4-induced liver fibrosis model[J]. Hepatology Communications, 2024, 8(4): 281-297.
[14] Samantha Lai, Simona Grassi, Vincenzo Nicola Talesa, Francesca Margheri, Lucia Magnelli. Endothelial Dysfunction in Chronic Kidney Disease: Highlighting the Role of FGF23 and Klotho[J]. International Journal of Molecular Sciences, 2024, 25(9): 5689-5689.
[15] Zhen Chen, Yingjie He, Yuchen Wu, Hui Chen, Xiaoyang Hou, Xiang Xiao, Yang Zhang, Jun Zou, Lei Pei, Jing Su, Chunlin Zhang, Xingdong Zhang, Yunsong Liu. Extracellular vesicles from mesenchymal stem cells promote chondrocyte proliferation and migration via miR-135b-5p-mediated modulation of FGF18 in osteoarthritis[J]. Stem Cell Research & Therapy, 2024, 15(1).
[16] Feng Wu, Yanan Zou, Yabin Xu, Chunjie Jiang, Haijun Bao, Ning Xie, Wenlong He. Bone marrow mesenchymal stem cells derived exosomes regulate VEGF/VEGFR2 signaling to promote lung injury repair[J]. Stem Cell Research & Therapy, 2024, 15(1).


Comments
Leave a Comment