Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
The difficulty in treating solid tumors lies largely in cancer-associated fibroblasts (CAFs) and the dense stroma they secrete, which act as key accomplices. They form a physical barrier that not only impedes drug penetration but also suppresses T cell infiltration, creating an immunologically "cold" environment. Fibroblast activation protein (FAP), which is highly expressed on the surface of CAFs, represents an ideal target to overcome this challenge.
On February 13, 2026, a research team led by Ellen Puré from the University of Pennsylvania, in collaboration with Nobel laureate Drew Weissman and Carl June, the "father of CAR-T therapy," published a landmark study in Cancer Immunology Research. They developed an anti-CD5 antibody-conjugated lipid nanoparticle (tLNP) encapsulating FAP-CAR mRNA. A single intravenous injection was sufficient to reprogram endogenous T cells into FAP-CAR-T cells in vivo.
In a pancreatic cancer model, this "in vivo CAR-T" technology demonstrated remarkable efficacy: it achieved an average tumor volume reduction of 74%, outperforming traditional ex vivo virally transduced CAR-T cells (48%). Importantly, this approach required no lymphodepleting preconditioning, carried no risk of genomic integration, and could rapidly eliminate FAP+ stromal cells, thereby remodeling the tumor microenvironment and allowing endogenous immune cells to infiltrate.
This breakthrough offers a safer, more convenient, and lower-cost "off-the-shelf" solution for solid tumor immunotherapy and represents a critical step forward in translating FAP-targeted therapies from concept to clinical application.
1. Background and Significance of FAP Research
2. Biological Characteristics, Structure, and Physiological Functions of FAP
3. Mechanisms and Signaling Pathways of FAP in Disease
Fibroblast activation protein (FAP) is a type II transmembrane serine protease with low expression under normal physiological conditions. However, its expression is significantly upregulated in various pathological contexts, particularly within the tumor microenvironment, garnering widespread attention [1-4]. Initially named for its expression on activated fibroblasts, FAP plays important roles in multiple diseases, especially fibrosis, inflammation, and cancer [4]. As a proteolytic enzyme, FAP cleaves proteins in the extracellular matrix (ECM), influencing cell-cell interactions and tissue remodeling, thereby promoting disease progression [1].The expression pattern of FAP makes it a potential biomarker and therapeutic target. Studies show that FAP is highly expressed on cancer-associated fibroblasts (CAFs) in over 90% of cancer types (e.g., breast cancer, pancreatic cancer, esophageal cancer, lung cancer), while being nearly absent in healthy adult tissues [2][3][5][6]. This pathology-specific expression endows FAP with great potential as a target for cancer diagnosis and treatment [6].
FAP possesses unique enzymatic activity, enabling it to specifically recognize and degrade proline-rich components of the ECM, such as collagen and gelatin [1][8]. In normal adult tissues, FAP expression is generally low, only upregulated during specific physiological processes like embryonic development and wound healing [1][4]. However, under various pathological conditions, especially in the tumor microenvironment, FAP expression is significantly increased and serves as a marker for CAFs [2][5].
FAP belongs to the dipeptidyl peptidase (DPP) family and is a proline-specific post-prolyl peptidase [10][8]. Its enzymatic activity allows it to cleave various ECM components, participating in tissue remodeling and pathological processes [1]. In normal tissues, FAP expression is low but transiently upregulated during processes like embryonic development and wound repair to support tissue repair [4].
TissuesAlthough FAP expression is low in normal adult tissues, it is temporarily upregulated during certain physiological processes, such as embryonic development and wound healing, where it plays important physiological roles [1][4]. By promoting fibroblast migration and proliferation, and participating in ECM remodeling, FAP supports tissue regeneration and repair [4]. This controlled expression and function highlight the key role of FAP in maintaining tissue homeostasis [4].
FAP's role in various diseases, particularly cancer, primarily involves influencing tumor cell biology through ECM remodeling and the regulation of intracellular signaling. Its expression in the tumor microenvironment promotes cancer cell invasion, metastasis, and therapy resistance. FAP not only degrades the ECM via its enzymatic activity but also regulates cell proliferation, survival, migration, and immune evasion by activating several key signaling pathways [10][11].
Within the tumor microenvironment, FAP affects tumor cell adhesion and migration by degrading ECM components. Research indicates that FAP activates the focal adhesion kinase (FAK) signaling pathway through interactions between integrin family members and the ECM, promoting tumor invasion and metastasis [10]. In non-small cell lung cancer (NSCLC), FAP further facilitates tumor metastasis by regulating the binding of integrins to ECM components [11]. Moreover, the interaction between FAP and integrins alters tumor cell adhesion strength and migration capacity, creating favorable conditions for invasion [11].
FAP regulates tumor cell proliferation, survival, and migration through multiple signaling pathways. For instance, in colorectal cancer (CRC), FAP promotes fibroblast proliferation and migration by activating the tumor necrosis factor receptor 2 (TNFR2)/Akt or ERK signaling pathways [17]. FAP also regulates cell proliferation and immune evasion through interaction with Yes-associated protein (YAP1). For example, in high-grade serous ovarian cancer (HGSOC), FAP+ CAFs inhibit CD8+ T cell cytotoxicity via a YAP1-dependent mechanism, reducing immune response and leading to poor patient prognosis [15].FAP is also closely associated with the transforming growth factor-beta (TGF-β) pathway. TGF-β promotes epithelial-mesenchymal transition (EMT) in bladder cancer cells via the FAP/VCAN axis, enhancing tumor cell invasiveness [16]. These findings reveal the important role of FAP in the tumor microenvironment, particularly its multiple mechanisms in tumor cell proliferation and immune evasion [11][17].
FAP-positive cancer-associated fibroblasts (CAFs) play multiple roles in the tumor microenvironment, crucially contributing to immune evasion, therapy resistance, and tumor metastasis. FAP+ CAFs induce radiotherapy resistance in tumor cells by secreting immunosuppressive factors like IL-8 [22]. Additionally, FAP+ CAFs alter ECM composition, preventing immune cell infiltration and forming a "cold" tumor microenvironment, further promoting immune evasion [20].FAP enhances the immunosuppressive function of CAFs through interactions with factors like CXCL12. For instance, in gastric cancer, FAP+ CAFs promote macrophage chemotaxis by secreting periostin (POSTN) and activate resistance to immune checkpoint blockers (ICB) via the Akt signaling pathway [21]. These studies indicate that FAP's role in the tumor microenvironment extends beyond ECM remodeling to include mechanisms like immune regulation and cell migration that drive malignant transformation [21].
The role of FAP in non-oncological diseases is gaining attention, particularly in fibrosis and chronic inflammatory diseases. For example, after myocardial ischemia-reperfusion injury (I/R), FAP expression is significantly upregulated in cardiac fibroblasts, promoting myocardial fibrosis [19]. Similarly, in liver fibrosis, FAP expression levels closely correlate with the activation of hepatic stellate cells, and FAP-α serves as an important diagnostic marker for fibrosis in clinical practice [18].
In oncology, the development of FAP-targeted radiopharmaceuticals is highly active, with numerous candidates advancing through clinical stages. This field is evolving from pure diagnostic or therapeutic applications towards a integrated "theranostic" approach. Concurrently, various other drug modalities are under investigation, including small molecule drugs, peptide-conjugated radionuclides, bispecific antibodies, and cell therapies. Selected investigational bispecific antibody and CAR-T programs are summarized in the table below.
| Edications | Mechanism of action | Type of medication | Under investigation indications (disease names) | Institution under Research | Highest Research Stage |
|---|---|---|---|---|---|
| RG7826 | 4-1BB Agonist | FAP Antagonist | Bispecific Antibody | Colorectal Cancer | Roche Holding AG | Phase 1/2 Clinical |
| RO-7567132 | FAP antagonists | TNFRSF3 agonists | Bispecific Antibody | Advanced Malignant Solid Tumors | Locally Advanced Malignant Solid Tumors | Metastatic Solid Tumors | Hoffmann-La Roche Ltd. | Clinical Phase 1 |
| RO-7300490 | CD40L stimulator | FAP regulator | Bispecific antibody | Locally advanced malignant solid tumor | Hoffmann-La Roche, Inc. | Clinical Phase 1 |
| BMS-986484 | CD40 inhibitor | FAP antagonist | Bispecific antibody | Advanced malignant solid tumors | Bristol Myers Squibb Co. | Clinical Phase 1 |
| SHR-7367 | CD40 Agonist | FAP Antagonist | Bispecific Antibody | Advanced cancer | Advanced malignant solid tumor | KRAS G12D mutation-positive solid tumor | Shanghai Hengrui Medicine Co., Ltd. | Clinical Phase 1 |
| BI-765179 | 4-1BB agonist | FAP modulator | Bispecific Antibody | Advanced Malignant Solid Tumors | Locally Advanced Malignant Solid Tumors, etc. | Boehringer Ingelheim International GmbH | Boehringer Ingelheim GmbH | Clinical Phase 1 |
| GEN1057 | DR4 Agonist | FAP Antagonist | Bispecific Antibody | Advanced Malignant Solid Tumors | Metastatic Solid Tumors | Solid Tumors | Genmab BV | Genmab, Inc. | Clinical Phase 1 |
| AGEN1721 | FAP regulator | TGF-β regulator | Bispecific antibody | Breast cancer | Colorectal cancer | Agenus, Inc. | Preclinical |
| Quadrivalent FAPxCD40 bispecific antibody (Hengrui Medicine) | CD40 agonist | FAP modulator | Bispecific antibody | Multivalent vaccine | Tumor | Jiangsu Hengrui Medicine Co., Ltd. | Preclinical |
| FAP-targeted CAR-T (Capstan Therapeutics) | FAP antagonist | CAR-T | Fibrosis | Capstan Therapeutics, Inc. | Preclinical |
| FAP CAR T cell therapy(Acuitas)FAP CAR T cell therapy (Acuitas) | FAP antagonist | CAR-T | Heart failure | Acuitas Therapeutics, Inc. | Preclinical |
| FAP-targeted CAR-T (Genethon) | FAP antagonist | CAR-T | Fibrosis | Duchenne muscular dystrophy | Genethon | Preclinical |
| Anti-FAP/TGF-PRII antibody (Merus) | FAP antagonist | TGFBR2 inhibitor | Bispecific antibody | Tumor | Incyte Corp. | Merus NVIncyte Corporation | Merus NV | Preclinical |
| M-300 (Mestag) | FAP modulators | TNFRSF3 agonists | Bispecific Antibody | Solid Tumor | Mestag Therapeutics Ltd. | Preclinical |
| Anti-OX40/FAP-α Bispecific Antibody (Roche) | FAP modulator | OX40 agonist | Bispecific antibody | Tumor | F. Hoffmann-La Roche Ltd. | Preclinical |
| OPTF01 | FAP antagonist | CAR-T | Glioblastoma | Optieum Biotechnologies, Inc. | Preclinical |
As a key target in the tumor microenvironment, FAP plays an important role in tumor invasion, metastasis, immune evasion, and drug resistance. Cusabio provides FAP recombinant proteins, antibodies, and ELISA kits to support the development of specific FAP-targeted drugs and explore their potential in cancer therapy.
● FAP Recombinant Protein
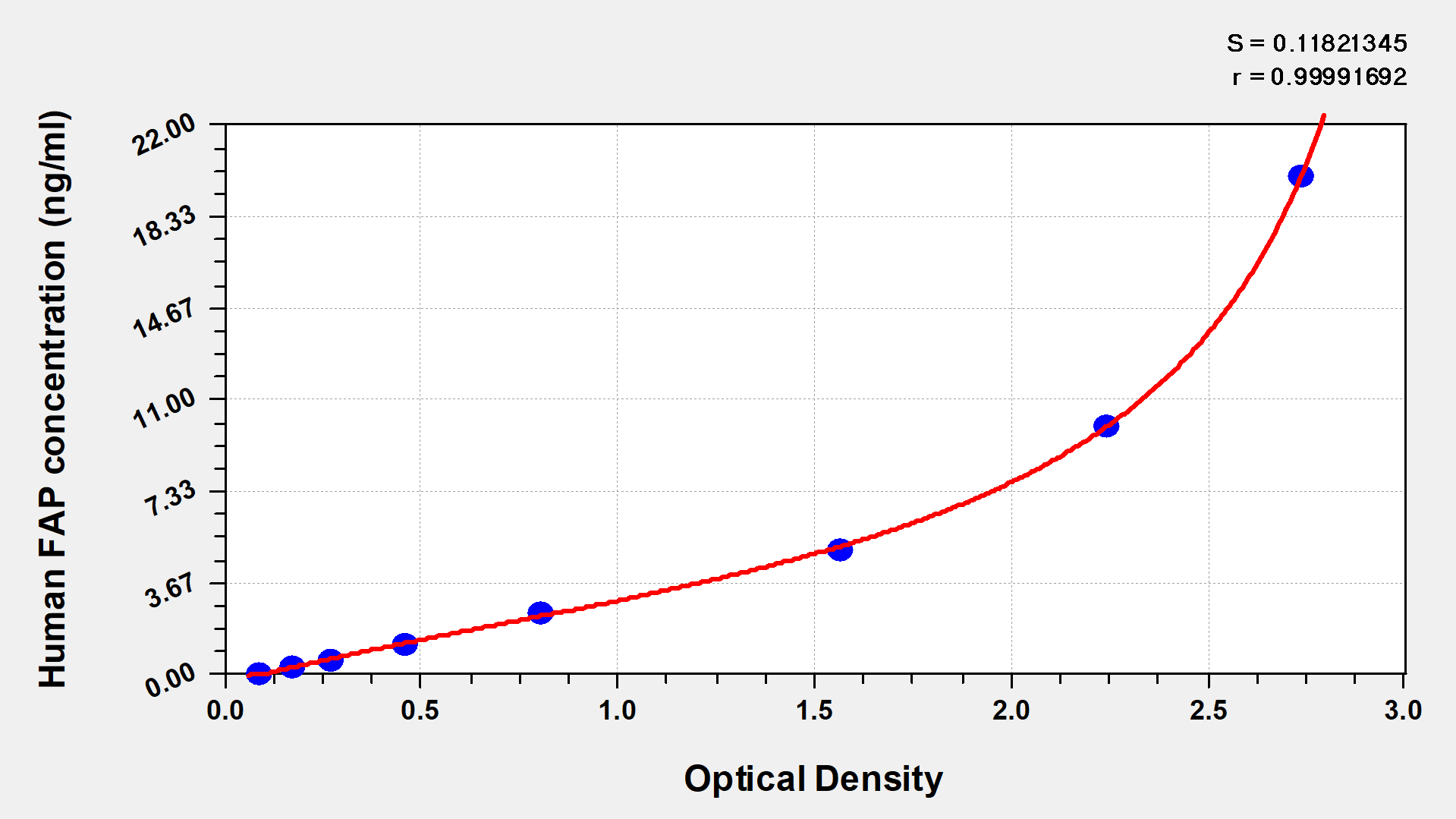
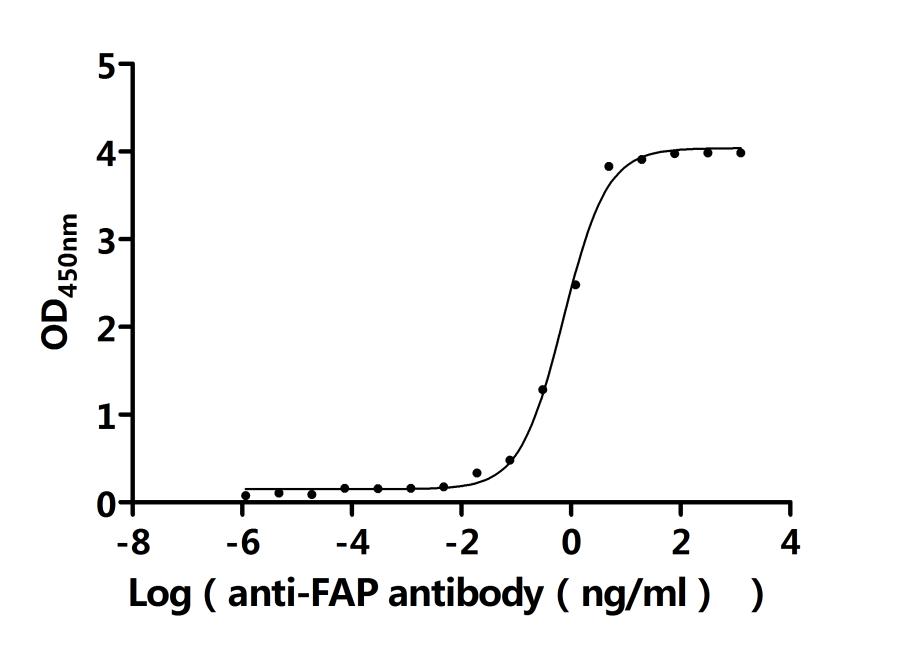
Recombinant Human Prolyl endopeptidase FAP (FAP), partial (Active); CSB-MP008424HU
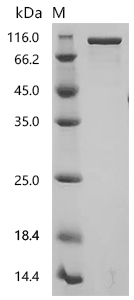
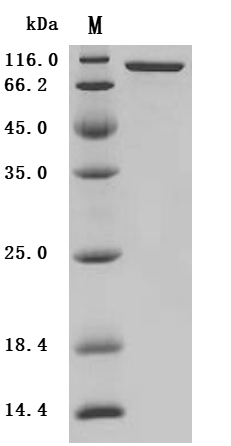
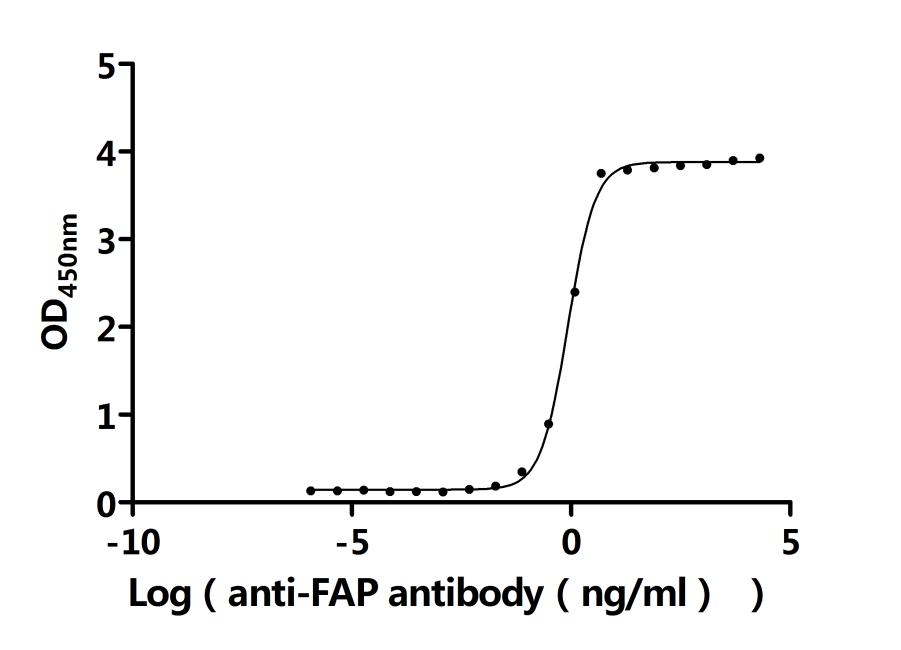
Recombinant Mouse Prolyl endopeptidase FAP (Fap), partial (Active); CSB-MP008424MO

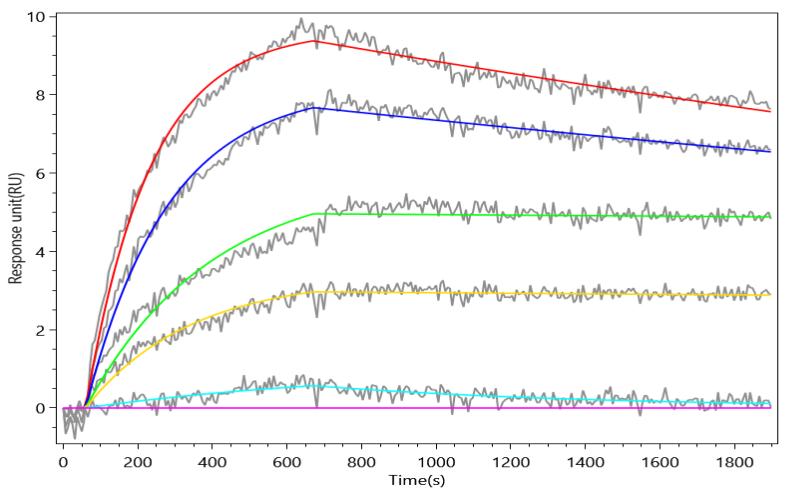
● FAP Antibody
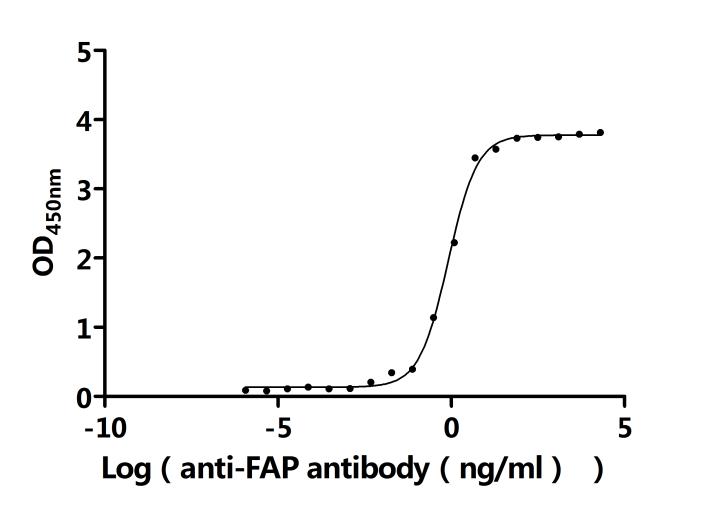
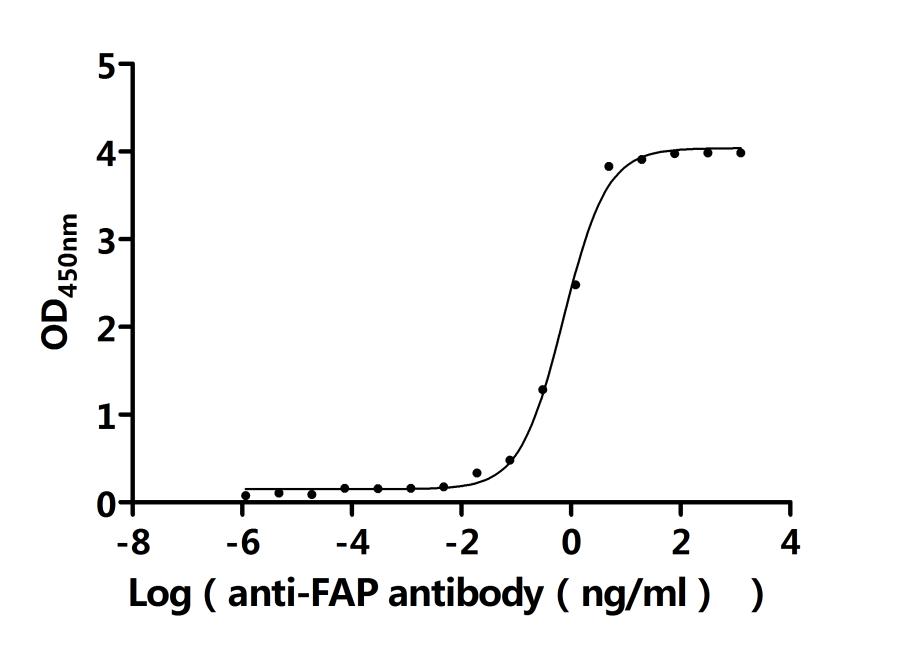
FAP Recombinant Monoclonal Antibody; CSB-RA008424MA1HU
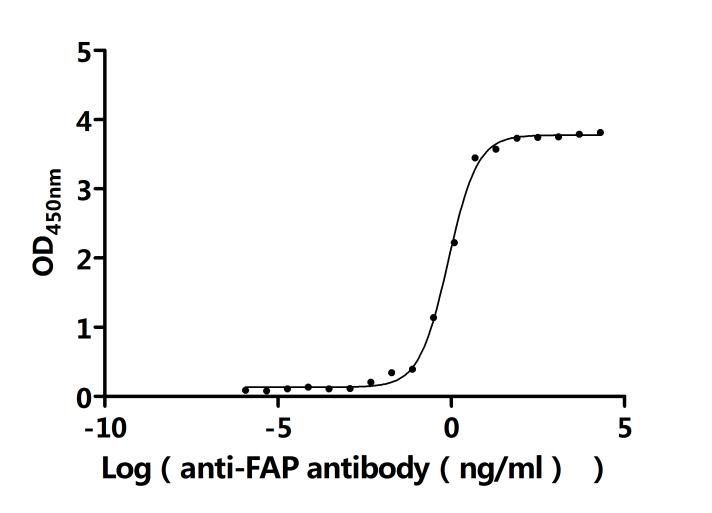
FAP Recombinant Monoclonal Antibody; CSB-RA008424MA2HU
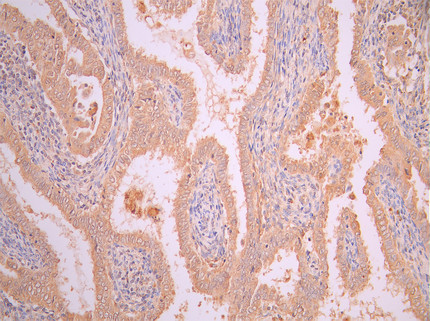
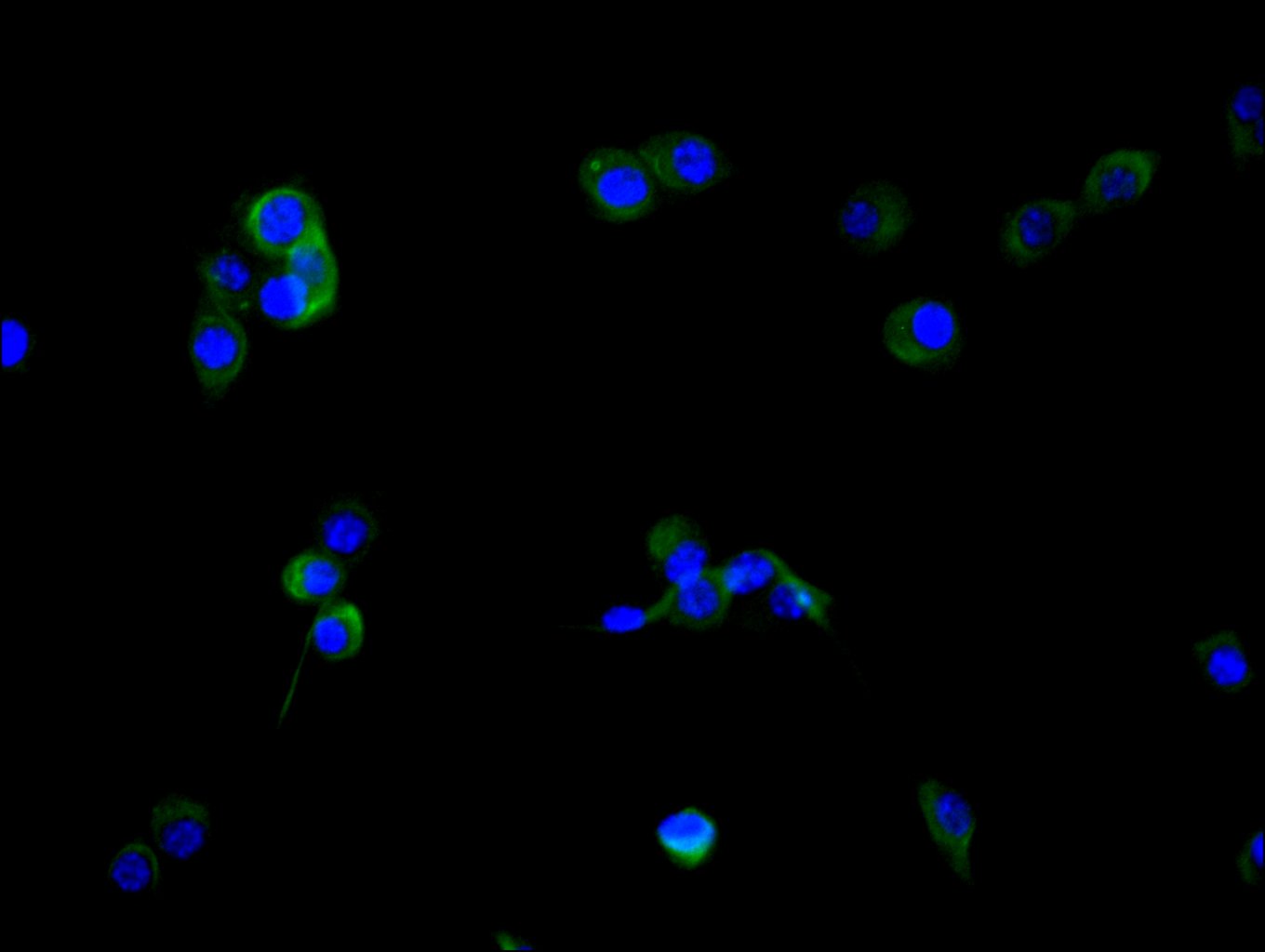
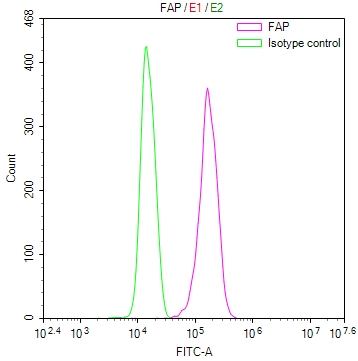
● FAP ELISA Kit
References
[1] Rasmus S Pedersen, M. Karsdal, N. Willumsen.(2024). Abstract 4287: Serological quantification of fibroblast activation protein (FAP) cleaved type III collagen: A biomarker for FAP activity.
[2] Sebastian Dziadek, A. Kraxner, Wei-Yi Cheng, Tai-Hsien Ou Yang, Mike Flores, Noah Theiss, T. Tsao, Emilia Andersson, S. V. Harring, Ann-Marie E. Bröske, Maurizio Ceppi, Volker Teichgräber, Jehad Charo.(2024). Comprehensive analysis of fibroblast activation protein expression across 23 tumor indications: insights for biomarker development in cancer immunotherapies.
[3] K. Hartmann, Merel van Gogh, P. C. Freitag, Florian Kast, Gabriela Nagy-Davidescu, L. Borsig, A. Plückthun.(2022). FAP-retargeted Ad5 enables in vivo gene delivery to stromal cells in the tumor microenvironment.
[4] Zihan Wang, Jinping Wang, Tianyi Lan, Liubo Zhang, Zeran Yan, N. Zhang, Yuan Xu, Qing-wen Tao.(2023). Role and mechanism of fibroblast-activated protein-α expression on the surface of fibroblast-like synoviocytes in rheumatoid arthritis.
[5] Spencer D. Lindeman, Jack Higgins.(2024). Abstract 6026: A novel lutetium-177 radioligand therapy targeting FAP has potent antitumor activity in xenograft cancer model.
[6] Ruchi Shah, Katherine A. Johnson, Anna E L Lippert, Sean G Kraus, Philip B. Emmerich, Cheri A Pasch, Wei Zhang, K. Matkowskyj, Aaron M LeBeau, D. Deming.(2024). Cancer-Associated Fibroblast Proteins as Potential Targets against Colorectal Cancers.
[7] Mengxin Xu, Pu Zhang, J. Ding, Junyi Chen, L. Huo, Zhibo Liu.(2021). Albumin Binder–Conjugated Fibroblast Activation Protein Inhibitor Radiopharmaceuticals for Cancer Therapy.
[8] Layne N. Raborn, Z. Michel, Michael T. Collins, Alison M Boyce, L. F. de Castro.(2024). Fibroblast Activation Protein Is Expressed by Altered Osteoprogenitors and Associated to Disease Burden in Fibrous Dysplasia.
[9] L. Loureiro, Lydia Hoffmann, Christin Neuber, Luise Rupp, C. Arndt, A. Kegler, M. Kubeil, Christoph E Hagemeyer, Holger Stephan, Marc Schmitz, A. Feldmann, M. Bachmann.(2023). Immunotheranostic target modules for imaging and navigation of UniCAR T-cells to strike FAP-expressing cells and the tumor microenvironment.
[10] Yentl Van Rymenant, Anke de Groot, Laura Dirkx, Emile Verhulst, Joni De Loose, Isabel Pintelon, Tias Verhezen, J. De Waele, Sofie Thys, O. De Wever, Muhammet Tanc, G. Caljon, Pieter Van der Veken, Ingrid De Meester.(2025). FAP on human NK cells: insights from NK cell activation and crosstalk with cancer-associated fibroblasts.
[11] Lirong Gao, Anqi Wang, Yuling Chen, Xin Cai, Yue Li, Jian Zhao, Yang Zhang, Weijie Zhang, Jianjie Zhu, Yuanyuan Zeng, Zeyi Liu, Jianyang Huang.(2023). FTO facilitates cancer metastasis by modifying the m6A level of FAP to induce integrin/FAK signaling in non-small cell lung cancer.
[12] Patrizio Ansalone.(2014). Electrostatic Affinities and Binding Kinetics among α2I Integrin Domains, Divalent Cations and 21-mer Collagen Fragment.
[13] Samuel Bell, Eugene M. Terentjev.(2016). Focal adhesion kinase - the reversible molecular mechanosensor.
[14] Min Li, Xue Cheng, Rong Rong, Yan-ping Gao, Xiuwu Tang, Youguo Chen.(2020). High expression of fibroblast activation protein (FAP) predicts poor outcome in high-grade serous ovarian cancer.
[15] Monika Licaj, R. Mhaidly, Y. Kieffer, H. Croizer, C. Bonneau, A. Meng, L. Djerroudi, Kevin Mujangi-Ebeka, H. Hocine, B. Bourachot, Ilaria Magagna, Renaud Leclère, Léa Guyonnet, Mylène Bohec, Coralie Guérin, S. Baulande, M. Kamal, C. le Tourneau, Fabrice Lécuru, Véronique Becette, Roman Rouzier, A. Vincent-Salomon, Géraldine Gentric, F. Mechta-Grigoriou.(2024). Residual ANTXR1+ myofibroblasts after chemotherapy inhibit anti-tumor immunity via YAP1 signaling pathway.
[16] Q. Ping, Chunhui Wang, Xin Cheng, Yiming Zhong, R. Yan, Meng Yang, Yunqiang Shi, Xiangmeng Li, Xiao Li, Wenwen Huang, Liqiong Wang, Xiaofang Bi, Libing Hu, Yang Yang, Yingbao Wang, R. Gong, Jun Tan, Rui Li, Hui Li, Jian Li, Wenju Wang, Ruhong Li.(2023). TGF-β1 dominates stromal fibroblast-mediated EMT via the FAP/VCAN axis in bladder cancer cells.
[17] Linlin Wang, Dong Yang, Jing Tian, Aiqin Gao, Yihang Shen, X. Ren, Xia Li, G. Jiang, Taotao Dong.(2017). Tumor necrosis factor receptor 2/AKT and ERK signaling pathways contribute to the switch from fibroblasts to CAFs by progranulin in microenvironment of colorectal cancer.
[18] Ruifang Li, Yifan Tai, Xinyan Zhang, Zhen Liu, Haipeng Si, Deling Kong, Lili Zhao, Jia Li, Adam C. Midgley.(2025). Tissue‐Microenvironment‐Responsive Self‐Assembling Peptide Nanoshells Boost Pirfenidone Efficacy in the Treatment of Liver Fibrosis.
[19] Jiawan Wang, Heng Du, Wanrun Xie, Jinmiao Bi, Hao Zhang, Xu Liu, Yuhan Wang, Shaolong Zhang, Anhua Lei, Chuting He, Hailong Yuan, Jiahe Zhang, Yujing Li, Pengfei Xu, Siqi Liu, Yanan Zhou, Jianghua Shen, Jingdong Wu, Yihong Cai, Chaofan Yang, Zeya Li, Y. Liang, Yang Zhao, Jin Zhang, Moshi Song.(2024). CAR-Macrophage Therapy Alleviates Myocardial Ischemia-Reperfusion Injury.
[20] Shipra Das, J. Valton, P. Duchateau, L. Poirot.(2023). Stromal depletion by TALEN-edited universal hypoimmunogenic FAP-CAR T cells enables infiltration and anti-tumor cytotoxicity of tumor antigen-targeted CAR-T immunotherapy.
[21] Tingting You, Huijing Tang, Wenjing Wu, Jin Gao, Xuechun Li, Ningning Li, Xiuxiu Xu, Jiazhang Xing, Hui Ge, Yi Xiao, Junchao Guo, Bin Wu, Xiaoyi Li, Liangrui Zhou, Lin Zhao, C. Bai, Qin Han, Zhao Sun, R. Zhao.(2023). POSTN Secretion by Extracellular Matrix Cancer-Associated Fibroblasts (eCAFs) Correlates with Poor ICB Response via Macrophage Chemotaxis Activation of Akt Signaling Pathway in Gastric Cancer.
[22] Weiqiang Huang, Longshan Zhang, Mi Yang, Xixi Wu, Xiaoqing Wang, Wenqi Huang, Lu Yuan, Hua Pan, Yin Wang, Zici Wang, Yuting Wu, Jihong Huang, Huazhen Liang, Shaoqun Li, Liwei Liao, Laiyu Liu, J. Guan.(2020). Cancer-associated fibroblasts promote the survival of irradiated nasopharyngeal carcinoma cells via the NF-κB pathway.


Comments
Leave a Comment