
Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
BTN3A1 (also known as CD277) is a key molecule for human Vγ9Vδ2 T cells to recognize phosphoantigens (pAg), and an important node connecting metabolic abnormality sensing, immune activation and therapeutic translation. Existing studies have shown that the B30.2 intracellular domain of BTN3A1 can selectively recognize pAg and induce conformational changes; subsequently, BTN3A1 forms a functionally related complex with BTN2A1, converting intracellular signals into a membrane surface structural state recognizable by Vγ9Vδ2 TCR. In addition to the classical γδ T cell activation function, BTN3A1 is also involved in NK cell regulation, T cell inhibition, and TBK1-IRF3-related innate immune signaling, indicating its obvious context-dependent and functional bidirectionality. Drug development targeting BTN3A1 has expanded to agonist antibodies, bispecific molecules, γδ TCR adapters, metabolic regulators and ADCs. Overall, BTN3A1 has clear biological and translational value, but its transmembrane signal transduction, complex dynamic remodeling, disease-specific functions and clinical stratification strategies still need to be further clarified.
1. Research Background and Biological Functions of BTN3A1 (CD277)
2. Structural Characteristics and Phosphoantigen (pAg) Recognition Mechanism of BTN3A1
3. Molecular Regulatory Mechanism of Vγ9Vδ2 T Cell Activation Mediated by BTN3A1
4. Immunomodulatory Bidirectionality and Multi-Pathway Functions of BTN3A1
5. Mechanism of BTN3A1 in Tumor and Autoimmune Diseases
6. Research Progress of BTN3A1-Targeted Drug Development and Cancer Immunotherapy Translation
7. BTN3A1 Research Tools: Recombinant Proteins and Antibody Products
BTN3A1 first attracted attention due to its core role in the recognition of phosphoantigens (pAg) by Vγ9Vδ2 T cells. Studies have found that the anti-CD277 antibody 20.1 can mimic Vγ9Vδ2 T cell activation caused by intracellular pAg accumulation, while 103.2 can block the activation of Vγ9Vδ2 T cells by target cells after pAg or aminobisphosphonate treatment; combined with siRNA knockdown and domain substitution experiments, the related function was finally localized to the BTN3A1 subtype [1]. Meanwhile, FRAP experiments showed that intracellular pAg accumulation reduces the membrane mobility of BTN3A1, and this change is closely related to subsequent T cell activation, providing direct evidence for the model of "metabolic abnormality - membrane protein dynamics change - γδ TCR recognition" [1]. In addition, ABCA1 mediates the efflux of representative pAg—isopentenyl pyrophosphate (IPP), and the interaction between apoA-I and BTN3A1 may be involved in this process, suggesting that soluble IPP may also participate in signal transmission between target cells and Vγ9Vδ2 T cells, but its specific recognition mode remains unclear [2].
With further research, the functional localization of BTN3A1 has been further refined. BTN2A1 has been shown to directly bind to the Vγ9 chain-related region, constituting the initial contact for γδ TCR recognition, while BTN3A1 regulates the spatial proximity to BTN2A1 through its intracellular domain, stabilizing cell contact and amplifying pAg-dependent signals [3]. In tumor immunity, the synergistic effect of BTN3A1 and BTN2A1 has also been confirmed to be exploitable by antibodies to enhance γδ T cell killing of tumor cells and improve the function of tumor-specific αβ T cells [4]. However, the functional interpretation of the CD277/BTN3 family still needs to be cautious, as commonly used CD277 antibodies cannot distinguish between BTN3 subtypes, and different cell types have different expression of BTN3A1, BTN3A2, and BTN3A3, which may affect the conclusions of early blocking or agonist experiments [5]. In inflammatory diseases, BTN3A1 is upregulated on the surface of monocytes in psoriasis patients and accompanied by abnormal Vγ9Vδ2 T cell function, also suggesting that BTN3A1 is not only a pAg sensing molecule, but also an important node of immune regulation [6].
The structural characteristics of BTN3A1 determine its function as an intracellular pAg sensing molecule. Its key region is the intracellular B30.2 (PRY/SPRY) domain, which forms a positively charged binding pocket that can accommodate negatively charged isoprenyl pyrophosphate molecules such as HMBPP and IPP; biochemical analysis, site-directed mutagenesis and binding experiments have shown that key residues such as H381 are critical for ligand recognition [7,8]. However, existing evidence suggests that simple "binding" is not sufficient to explain the antigen sensing function of BTN3A1. NMR, crystallography and molecular dynamics studies further indicate that truly antigenic pAg not only enters the B30.2 pocket, but also induces a global conformational change of the domain, and this allosteric response better explains the functional activation of γδ T cells than simple binding [9,10]. In addition, some ligands with similar binding affinity cannot effectively induce T cell responses, indicating that "binding ability" and "activation ability" are not completely equivalent [7]. Further structural studies have also found that regions adjacent to B30.2 and the juxtamembrane segment may participate in dimer formation and transmit intracellular conformational changes to the membrane region and extracellular region, thereby providing a basis for subsequent signal amplification [10-12].
How intracellular recognition is converted into an extracellular recognizable structure is one of the core questions in BTN3A1 research. Biochemical reconstitution, ITC and live-cell BRET experiments showed that pAg can induce the interaction between the intracellular regions of BTN3A1 and BTN2A1, and the integrity of the BTN2A1 juxtamembrane segment is critical for this process and subsequent IFNγ responses [13]. Recent cryo-EM studies further suggest that pAg-induced BTN3A1/BTN2A1 complex remodeling can promote BTN molecules to form a conformational architecture suitable for Vγ9Vδ2 TCR recognition [14]. Meanwhile, antibody blocking and engineered Fab spacing experiments also suggest that signal triggering does not depend on simple aggregation, but on precise geometric configuration and epitope exposure state [15]. Therefore, the more reasonable current understanding is that BTN3A1 is not just a passive "ligand binder", but a conformational sensor that translates intracellular metabolic abnormalities into recognizable signals on the membrane surface.
During Vγ9Vδ2 T cell recognition, pAg is not just a soluble metabolite, but more like a "molecular glue". It is first sensed by the intracellular domain of BTN3A1, then promotes the formation of a heterologous complex between BTN3A1 and BTN2A1, and drives signal transmission from the inside to the outside of the cell. Structural and kinetic studies have shown that pAg can be located at the key position of the BTN3A1–BTN2A1 complex interface, stabilizing the intracellular contact between the two and promoting dynamic changes conducive to the appearance of outward conformations [16]. Genetic and cytological studies have further confirmed that BTN2A1 is an indispensable component in the pAg sensing pathway and can directly contact the Vγ9-related region, so the BTN3A1–BTN2A1 complex should be regarded as a functional whole rather than two independent molecules [17]. However, most current evidence still comes from truncated constructs or in vitro systems, and there is still a lack of direct observation of the dynamic behavior of full-length membrane proteins in the natural membrane environment [18].
Studies have shown that BTN2A1 and BTN3A1 together form the composite ligand required for Vγ9Vδ2 TCR recognition: BTN2A1 laterally contacts the TCR, while BTN3A1 provides another recognition epitope, and the complex formed by the two and its dynamic state determine the recognition efficiency [19]. From the perspective of the signal pathway, after pAg binds to the intracellular B30.2 (PRY/SPRY) domain of BTN3A1, it can trigger extracellular conformational changes through the "inside-out" method; this stimulation is highly consistent with the activation induced by anti-BTN3A antibodies in downstream phosphorylation events such as ZAP70, PLCγ2, Akt, NFκB, p38 and Erk [20,21]. In addition, the protein stability, membrane localization and subcellular trafficking of BTN3A1 also affect this process. Related studies suggest that transporters such as ABCG2 are associated with BTN3A-dependent activation, while BTN3A2 may regulate the trafficking and optimal activity of BTN3A1 [22,23]. Therefore, BTN3A1-mediated Vγ9Vδ2 T cell activation is not a single ligand binding event, but a process jointly regulated by complex assembly, membrane localization and intracellular delivery [19-23].
BTN3A1 does not only serve Vγ9Vδ2 T cell recognition. Studies have found that pAg-stimulated γδ T cells can inhibit NK cell responses to "missing-self" targets in a BTN3A1-dependent, cell contact-dependent manner, accompanied by downregulation of NKG2D and NKp44, suggesting that BTN3A1 may be involved in the negative regulation between innate immune effector cells [24]. On the other hand, LSECtin binding to BTN3A1 inhibits human T cell proliferation and inflammatory factor production, and this inhibition can be partially reversed by anti-BTN3A1 antibodies, indicating that BTN3A1 can also act as a negative signal receptor on the cell surface [25].
In contrast, BTN3A1 also has a positive role in innate immunity. Studies have shown that BTN3A1 can form a complex with TBK1, and under nucleic acid stimulation, it is relocated to the perinuclear region through MAP4-controlled dynein-dependent transport, promoting the interaction between TBK1 and IRF3 and enhancing IFN-β production; knockdown of BTN3A1 impairs the type I interferon response of cells to cytoplasmic DNA or RNA [26]. This indicates that BTN3A1 has obvious functional bidirectionality: it can participate in inhibitory cell-cell contact signals and also act as an intracellular scaffold to promote antiviral pathway activation. Therefore, any future therapeutic design targeting BTN3A1 must fully consider its domain division, stimulation context and cell type differences.
In tumor immunity, BTN3A1 has a significant dual role. On the one hand, the synergy between BTN3A1 and BTN2A1, as well as related juxtamembrane regulatory events, are important foundations for Vγ9Vδ2 T cells to recognize and eliminate early abnormal cells; proteomics studies also suggest that factors such as PHLDB2, SYNJ2, and CARMIL1 can regulate the surface dynamics of BTN2A1/BTN3A1, thereby affecting anti-tumor surveillance [27]. On the other hand, in models such as cervical cancer, high expression of BTN3A1 in tumor cells can directly drive Vδ2 T cell exhaustion, induce upregulation of NR4A2/3 and reduce effector molecule secretion; meanwhile, IFN-γ produced by Vδ2 T cells can in turn promote upregulation of BTN3A1 and PD-L1 in tumor cells, forming a negative feedback immunosuppressive loop [28]. Therefore, although antibodies targeting BTN3A1 have shown potential to enhance anti-tumor responses in models, how to balance "promoting activation" and "preventing exhaustion" remains a key issue in translation [29].
The relationship between BTN3A1 expression and prognosis is not consistent across different cancer types. In some breast cancer cohorts, high BTN3A1 expression is associated with better survival outcomes and higher immune infiltration [30]; while in gliomas, high BTN3A1 expression is more associated with immunosuppressive characteristics and poor prognosis [31]. This indicates that a single expression level is not sufficient as a stable prognostic or therapeutic biomarker, and must be comprehensively judged in combination with cell type, partner molecules and signal modification status.
In addition to tumors, BTN3A1 may also be involved in autoimmune diseases. Mendelian randomization, colocalization and multi-omics analysis have shown that BTN3A1 plasma protein levels are significantly associated with IgA nephropathy risk, and it has also been identified as a candidate related gene in Sjögren's syndrome studies [32,33]. However, such genetic and multi-omics evidence still requires further tissue and functional verification to clarify its true causal role and drug development value.
Drug development targeting the BTN3A1 target has entered the early clinical validation stage, and the fastest-progressing candidate drug (ICT-01) globally has advanced to Phase 1/2 clinical trials in acute myeloid leukemia and advanced solid tumors. Overall, the target has attracted the layout of many institutions at home and abroad, with drug forms covering monoclonal antibodies, small molecules and TCR fusion proteins, etc., with indications mainly focusing on tumors, while expanding to fields such as infections and autoimmune diseases, but the vast majority of projects are still in the preclinical or drug discovery stage.
| Medications | Target | Types of medications | Indications under investigation | Research institutions | Highest R&D stage |
|---|---|---|---|---|---|
| ICT-01 | BTN3A1 | Monoclonal antibodies | Acute Myeloid Leukemia | Acute Lymphoblastic Leukemia | Advanced Cancer | advanced malignant solid tumors, etc | Imcheck Therapeutics SAS | Clinical phase 1/2 |
| ICT-41 | BTN3A1 | Monoclonal antibodies | Bacterial infections | Viral infection | Imcheck Therapeutics SAS | Preclinical |
| CD20-targeted GADLEN(Shattuck Labs) | BTN2A1 x BTN3A1 x CD20 x CD3 | TCR Fusion Protein | T Cell Binder | Antibody fusion protein | tumors | Shattuck Labs, Inc. | Preclinical |
| CD19GADLEN | BTN2A1 x BTN3A1 x CD19 | Antibody fusion protein | Lymphoma | Shattuck Labs, Inc. | Preclinical |
| BTN3A1 ligand Compound 26b(University of Iowa) | BTN3A1 | Chemicals | tumors | University of Iowa | Preclinical |
| B7-H3 GADLEN | BTN2A1 x BTN3A1 x CD20 x CD276 | Antibody fusion protein | tumors | Shattuck Labs, Inc. | Preclinical |
| BSI-093 | BTN3A1 | Monoclonal antibodies | Biliary tract cancer | Esophageal cancer | Squamous cell carcinoma of the head and neck | Biotechnology (Nanjing) Co., Ltd | Preclinical |
| POM2-C-HMBP | BTN3A1 | Small molecule chemicals | tumors | University of Iowa | University of Connecticut | Drug discovery |
| WO2024146627 | BTN3A1 | Small molecule chemicals | Autoimmune Diseases | Cardiovascular Disease | Infection | tumors | Beijing Qinghui Liannuo Biotechnology Co., Ltd | Drug discovery |
| Compound 6e(MIT-World Peace University) | BTN3A1 | Small molecule chemicals | - | University of Connecticut | Drug discovery |
| US20230285527 | BTN3A1 | Monoclonal antibodies | Immune system disorders | Université d'Aix-Marseille | Nantes Universite | Centre National de la Recherche Scientifique | Institut Jean Paoli & Irene Calmettes | Drug discovery |
| CN117295505 | BTN3A1 | Cell therapy | Immune System Disorders | tumors | The Regents of the University of California | The J. David Gladstone Institutes | Drug discovery |
| WO2024146629 | BTN3A1 | Small molecule chemicals | Autoimmune Diseases | Cardiovascular Disease | Infection | tumors | Beijing Qinghui Liannuo Biotechnology Co., Ltd | Drug discovery |
(Data as of March 12, 2026, source: Synapse)
Currently, BTN3A1 has evolved from a family member annotation molecule to a core node explaining the mechanism of phosphoantigen (pAg) recognition by human Vγ9Vδ2 T cells. Cusabio provides BTN3A1 recombinant protein and antibody products to support your mechanism research and targeted drug development.
● BTN3A1 Recombinant Proteins
● BTN3A1 Antibodies
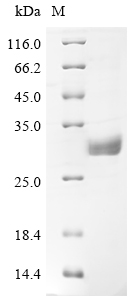
BTN3A1 Recombinant Monoclonal Antibody; CSB-RA002873MA2HU
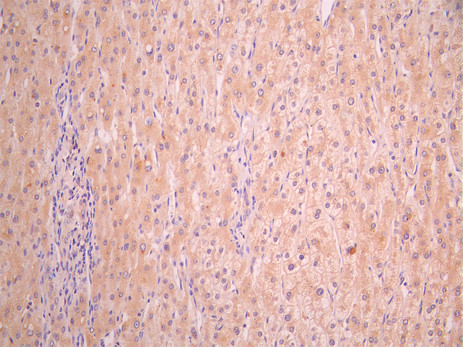
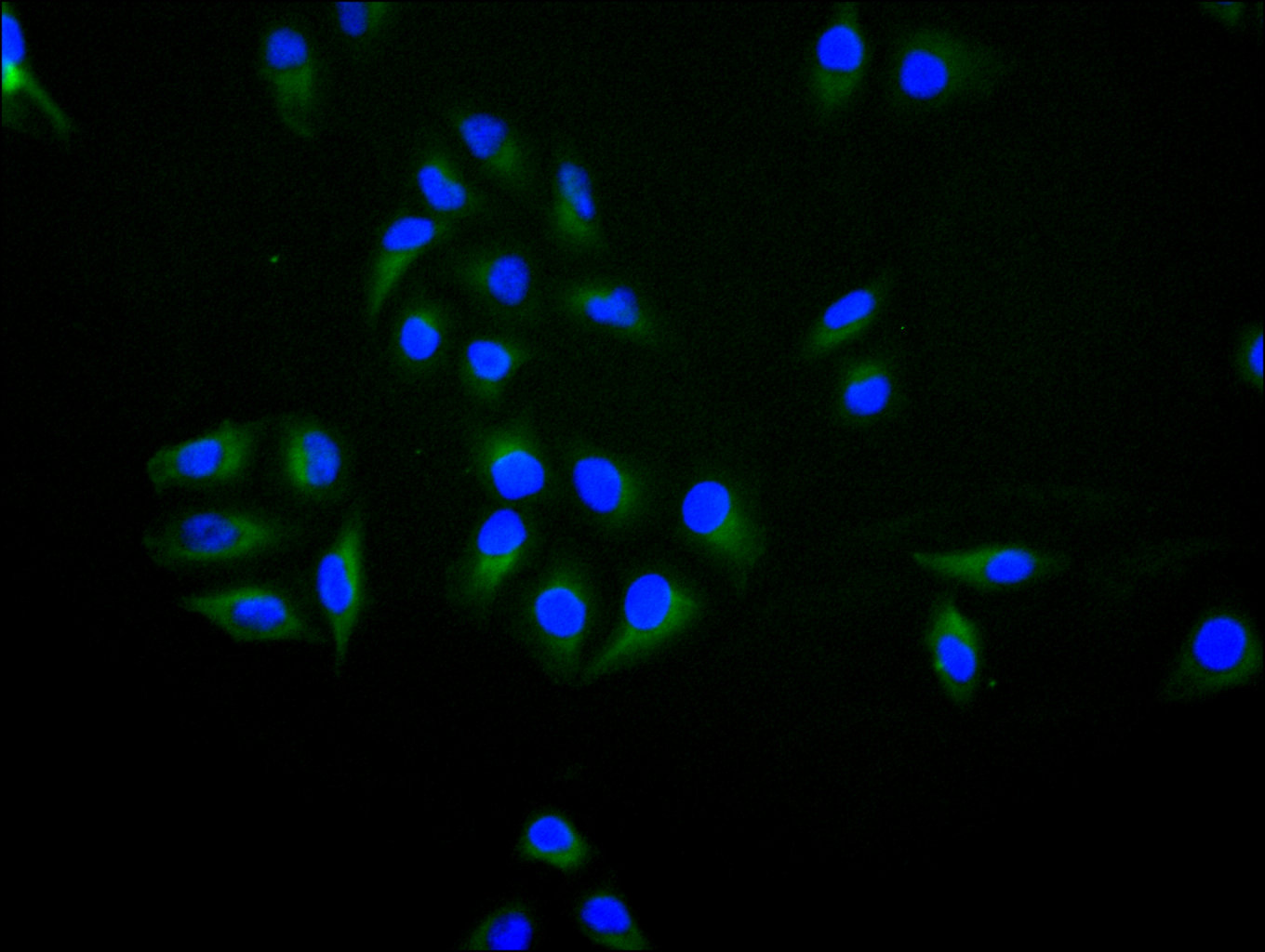
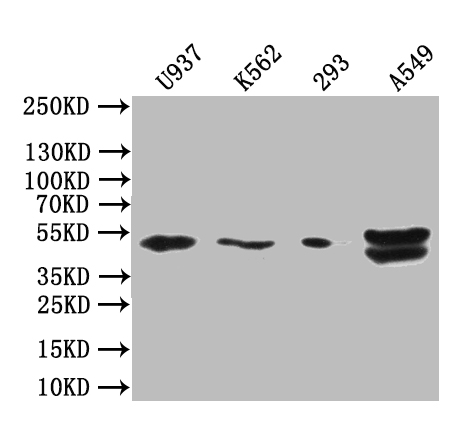
BTN3A1 Antibody; CSB-PA002873LA01HU
References
[1] Harly C, Guillaume Y, Nedellec S, et al. Key implication of CD277/butyrophilin-3 (BTN3A) in cellular stress sensing by a major human γδ T-cell subset. Blood. 2012;120(11):2269-2279.
[2] Riganti C, Castella B, Massaia M. ABCA1, apoA-I, and BTN3A1: A Legitimate Ménage à Trois in Dendritic Cells. Front Immunol. 2018;9:2802.
[3] Vyborova A, Beringer DX, Fasci D, et al. γ9δ2T cell diversity and the receptor interface with tumor cells. Nat Commun. 2020;11(1):6296.
[4] Kelly PN. BTN3A1 governs antitumor responses. Science. 2020;369(6503):504-505.
[5] Messal N, Mamessier E, Sylvain A, et al. Differential role for CD277 as a co‐regulator of the immune signal in T and NK cells. Eur J Immunol. 2011;41(11):3234-3244.
[6] Zhou J, Zhang J, Tao L, et al. Up-regulation of BTN3A1 on CD14+ monocytes in psoriasis patients contributes to Vγ9Vδ2 T cell activation. J Dermatol Sci. 2022;108(3):179-187.
[7] Poe MM, Agabiti SS, Liu C, et al. Probing the Ligand-Binding Pocket of BTN3A1. ACS Chem Biol. 2019;14(12):2681-2691.
[8] Wang H, Morita CT. Sensor Function for Butyrophilin 3A1 in Prenyl Pyrophosphate Stimulation of Human Vγ2Vδ2 T Cells. J Immunol. 2015;195(5):2359-2368.
[9] Salim M, Knowles TJ, Baker AT, et al. BTN3A1 Discriminates γδ T Cell Phosphoantigens from Nonantigenic Small Molecules via a Conformational Sensor in Its B30.2 Domain. J Biol Chem. 2017;292(16):6662-6673.
[10] Gu S, Sachleben JR, Boughter CT, et al. Phosphoantigen-induced conformational change of butyrophilin 3A1 (BTN3A1) and its implication on Vγ9Vδ2 T cell activation. Proc Natl Acad Sci U S A. 2017;114(30):E6119-E6128.
[11] Peigné CM, Léger A, Gesnel MC, et al. The Juxtamembrane Domain of Butyrophilin BTN3A1 Controls Phosphoantigen-Mediated Activation of Human Vγ9Vδ2 T Cells. Cell Rep. 2017;19(13):2773-2785.
[12] Nguyen K, Hsiao CHC, Jin Y, Wiemer AJ, Vinogradova O. Investigation of structural and dynamic properties of the Butyrophilin BTN3A1/BTN2A1 cytoplasmic complex by NMR spectroscopy. J Biomol NMR. 2025;79(1):1-14.
[13] Hsiao CHC, Nguyen K, Jin Y, Vinogradova O, Wiemer AJ. Ligand-induced interactions between butyrophilin 2A1 and 3A1 internal domains in the HMBPP receptor complex. J Biol Chem. 2022;298(12):102684.
[14] Zhang M, Wang Y, Cai N, et al. Cryo-EM structural insights into Vγ9Vδ2 TCR activation via multiple butyrophilins. Cell Res. 2024;34(2):128-141.
[15] Ramesh A, Roy S, Slezak T, et al. Mapping the extracellular molecular architecture of the pAg-signaling complex with α-Butyrophilin antibodies. Cell Rep. 2025;48(3):113568.
[16] Yuan L, Ma X, Yang Y, et al. Phosphoantigens glue butyrophilin 3A1 and 2A1 to activate Vγ9Vδ2 T cells. Nature. 2023;621(7980):649-656.
[17] Rigau M, Ostrouska S, Fulford TS, et al. Butyrophilin 2A1 is essential for phosphoantigen reactivity by γδ T cells. Science. 2020;367(6477):eaay5516.
[18] Yuan L, Ma X, Yang Y, et al. Phosphoantigens are Molecular Glues that Promote Butyrophilin 3A1/2A1 Association Leading to Vγ9Vδ2 T Cell Activation. bioRxiv. 2022:2022.07.25.501389.
[19] Fulford TS, Soliman C, Castle RG, et al. Vγ9Vδ2 T cells recognize butyrophilin 2A1 and 3A1 heteromers. Nat Immunol. 2023;24(3):424-435.
[20] Gu S, Nawrocka WI, Adams EJ. Sensing of Pyrophosphate Metabolites by Vγ9Vδ2 T Cells. J Immunol. 2015;195(5):2049-2056.
[21] Decaup E, Duault C, Bezombes C, et al. Phosphoantigens and butyrophilin 3A1 induce similar intracellular activation signaling in human TCRVγ9+ γδ T lymphocytes. Eur J Immunol. 2014;44(11):3323-3333.
[22] Rhodes DA, Chen HC, Williamson JC, et al. Regulation of Human γδ T Cells by BTN3A1 Protein Stability and ATP-Binding Cassette Transporters. Cell Rep. 2018;24(6):1548-1560.e6.
[23] Vantourout P, Laing A, Woodward MJ, et al. Heteromeric interactions regulate butyrophilin (BTN) and BTN-like molecules governing γδ T cell biology. Proc Natl Acad Sci U S A. 2018;115(5):1039-1044.
[24] Walwyn-Brown K, Pugh J, Cocker ATH, et al. Phosphoantigen-Stimulated γδ T Cells Suppress Natural Killer–Cell Responses to Missing-Self. Front Immunol. 2022;13:864704.
[25] Wang J, Manick B, Renelt M, et al. LSECtin interacts with BTN3A1 to inhibit T cell activation. Mol Immunol. 2020;126:120-128.
[26] Seo M, Lee SO, Kim JH, et al. MAP4-regulated dynein-dependent trafficking of BTN3A1 controls the TBK1-IRF3 signaling axis. Proc Natl Acad Sci U S A. 2016;113(29):8266-8271.
[27] Cleven A, Meringa AD, Brázda P, et al. Sensitivity to Vγ9Vδ2TCR T cells is imprinted after single mutations during early oncogenesis. Cell. 2024;187(1):182-198.e24.
[28] Liu J, Wu M, Yang Y, et al. BTN3A1 expressed in cervical cancer cells promotes Vγ9Vδ2 T cells exhaustion through upregulating transcription factors NR4A2/3 downstream of TCR signaling. J Immunother Cancer. 2024;12(4):e008795.
[29] Payne KK, Mine JA, Biswas S, et al. BTN3A1 governs antitumor responses by coordinating αβ and γδ T cells. Science. 2020;369(6503):593-599.
[30] Ren H, Li S, Liu X, et al. Multi-omics analysis of the expression and prognostic value of the butyrophilins in breast cancer. Cancer Cell Int. 2021;21(1):619.
[31] Kone AS, Ghouzlani A, Qandouci A, et al. High expression of BTN3A1 is associated with clinical and immunological characteristics and predicts a poor prognosis in advanced human gliomas. J Neurooncol. 2024;166(2):387-398.
[32] Xu X, Miao C, Yang S, et al. Investigating potential drug targets for IgA nephropathy and membranous nephropathy through multi-queue plasma protein analysis: a Mendelian randomization study based on SMR and co-localization analysis. Front Immunol. 2024;15:1372489.
[33] Bai Y, Wang J, Feng X, et al. Identification of drug targets for Sjögren's syndrome: multi-omics Mendelian randomization and colocalization analyses. Front Immunol. 2024;15:1357177.
[34] Oberg HH, Deseke M, Krohn S, et al. Tumor-restricted activation of Vγ9Vδ2 T cells via bispecific Evobodies: a novel strategy for safe and potent immunotherapy in ovarian cancer. Cancer Immunol Immunother. 2025;74(3):689-702.


Comments
Leave a Comment