Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
CD4 is a key molecule on the surface of helper T cells and plays a central role in adaptive immune responses. This article systematically reviews the structure and function of CD4, the differentiation and regulatory mechanisms of CD4⁺ T cell subsets, with a focus on the roles of signaling pathways such as TCR, JAK/STAT, and TGF-β/SMAD in immune regulation. It also summarizes the research progress of CD4 in diseases including HIV/AIDS, autoimmune diseases, and cancer, as well as the current landscape of drug development targeting CD4.
1. Introduction: The Core Position and Research Value of CD4 in the Immune System
2. Molecular Characteristics and Basic Functions of CD4
3. CD4+ T Cell Biology: Subsets, Differentiation and Regulatory Mechanisms
4. Key Signaling Pathways Regulating CD4+ T Cell Function
5. The Role of CD4+ T Cells in the Occurrence and Development of Diseases
CD4 is a key transmembrane glycoprotein in the adaptive immune system, mainly expressed on the surface of helper T cells (CD4+ T cells), monocytes, macrophages and dendritic cells. Among them, on CD4+ T cells, CD4 acts as a coreceptor for the T cell receptor (TCR), and by binding to major histocompatibility complex class II (MHC-II) molecules, it cooperatively mediates T cell activation, making it an important hub for the initiation and regulation of adaptive immune responses.
CD4+ T cells have significant functional diversity and can differentiate into multiple subsets such as Th1, Th2, Th17 and Treg, playing irreplaceable roles in immune assistance, inflammatory regulation and maintenance of immune tolerance [1]. In recent years, with the development of imaging technology, single-cell analysis and computational models, researchers' understanding of CD4 molecules has continued to deepen. For example, super-resolution imaging studies have revealed the nanoscale distribution of CD4 on the plasma membrane of resting T cells and its characteristics affected by palmitoylation [2]; tumor immune microenvironment studies have shown that automatic identification and classification of CD4+ T cells contributes to prognostic evaluation and treatment response analysis [3]; artificial intelligence-driven imaging analysis also shows potential in distinguishing CD4 and CD8 cells [4].
At the same time, drug development targeting CD4 continues to advance. Taking the humanized anti-CD4 monoclonal antibody Zanolimumab as an example, it can inhibit CD4+ T cells by inhibiting TCR signal transduction, inducing Fc-dependent effector mechanisms and down-regulating cell surface CD4 expression, and is currently used in related research on skin and lymph node T cell lymphoma [5]. These advances indicate that CD4 is not only a key molecule for understanding immune regulation mechanisms, but also has important disease research and drug development value.
CD4 is usually a single-chain transmembrane glycoprotein, composed of four immunoglobulin-like extracellular domains (D1-D4), a transmembrane region and a shorter intracellular region. The extracellular D1 and D2 domains are mainly involved in binding to MHC class II molecules, while the intracellular region is closely associated with the Lck tyrosine kinase, thereby participating in the initiation and regulation of TCR signaling.
In addition to classical CD4+ T cells, CD4 is also expressed on the surface of immune cells such as macrophages, dendritic cells and monocytes, so its function is not limited to T cell activation, but is also closely related to processes such as antigen presentation, viral infection and immune microenvironment regulation. In recent years, quantitative imaging methods based on TIRF and SOFI have improved the resolution of nanoscale distribution of CD4 on the membrane surface, further confirming the importance of palmitoylation for the distribution pattern of CD4 [2]. In addition, artificial intelligence-assisted label-free imaging technology also suggests that CD4 has high application potential as a cell surface marker in cell recognition and classification [4].
In T cell-mediated immune responses, the core role of CD4 is to assist TCR in recognizing antigen peptides presented by MHC class II molecules on the surface of antigen-presenting cells. CD4 enhances the stability of the TCR-pMHC complex by binding to MHC II, thereby improving the efficiency and specificity of T cell antigen recognition.
In addition to stabilizing the recognition complex, CD4 can also recruit and regulate the activity of p56lck through the intracellular region. Activated p56lck can phosphorylate the ITAM motif of the CD3 chain in the TCR complex, thereby initiating the early signal cascade of T cells. It is worth noting that studies on anti-CD4 antibodies suggest that CD4 is not only involved in activation, but also related to inhibitory regulation. Studies have shown that Zanolimumab can rapidly inhibit early TCR signaling and activate CD4-associated p56lck, which suggests that it may change the coupling state of p56lck and the TCR complex, and instead activate inhibitory signaling molecules such as Dok-1 and SHIP-1, thereby achieving inhibition of T cell function [5]. This indicates that CD4 is not just a simple coreceptor, but also an important node connecting recognition, kinase recruitment and signal regulation.
CD4+ T cells are highly heterogeneous and can differentiate into different subsets according to transcription factor expression and cytokine secretion profiles. Classical subsets include Th1, Th2, Th17 and Treg:
In addition to classical Th subsets, Tfh and CD4 CTL are also important types that have received high attention in recent years:
The differentiation of CD4+ T cells is jointly regulated by antigen strength, cytokine environment and transcription factor network:
At the intracellular regulatory level, transcription factors such as Bcl6, Maf, ThPOK, Runx3, and Eomes are involved in shaping the fate and function of different CD4+ T cell subsets [13],[18-19]. These factors antagonize or cooperate with each other, enabling CD4+ T cells to dynamically remodel according to different microenvironments such as infection, inflammation or tumor, thereby reflecting strong plasticity. This plasticity is the basis for the flexibility of immune response, and also increases the complexity of disease research and therapeutic intervention.
After activation, CD4+ T cells need to undergo rapid proliferation and meet effector functional requirements through metabolic reprogramming. Their proliferation and survival are supported by cytokines such as IL-7, and are also regulated by metabolic pathways, mitochondrial status and intracellular energy balance [27]. Different subsets have differences in metabolic patterns. For example, inflammatory effector T cells usually prefer glycolysis, while regulatory T cells rely more on oxidative metabolism. Metabolic state not only affects cell survival, but also determines their differentiation direction and functional output. Therefore, metabolic regulation has become an important entry point for understanding the homeostasis and disease association of CD4+ T cells.
TCR proximal signaling is the starting point of CD4+ T cell activation. CD4 assists MHC II recognition and recruits Lck, promotes ITAM phosphorylation of the CD3 chain, and then activates downstream signaling molecules such as ZAP-70. Subsequently, calcium signal and calcineurin-NFAT pathway are activated, driving effector gene expression and regulating T cell differentiation and functional output. This pathway has a fundamental role in the early fate determination of T cells.
The JAK/STAT pathway is one of the core pathways for cytokine to regulate CD4+ T cell differentiation. Different cytokines can promote the formation of Th1, Th17 or Treg subsets by activating different STAT members. Studies have shown that some natural products or immunomodulatory factors can change the balance between Th1/Th17 and Treg by inhibiting the activity of STAT1, STAT3 or STAT5, thereby exerting therapeutic potential in inflammatory and autoimmune diseases [9-10].
The TGF-β/SMAD pathway has an important position in Treg formation, immune tolerance maintenance and inflammatory regulation; Notch signaling plays an auxiliary regulatory role in T cell development, differentiation and functional shaping. These two pathways do not exist in isolation, but interact with pathways such as JAK/STAT to jointly participate in the fate regulation of CD4+ T cells.
The cGAS/STING pathway was originally mainly considered to be involved in cytosolic DNA sensing and innate immune activation, but recent studies suggest that this pathway also affects the tumor immune microenvironment and T cell functional status. Events such as metabolic stress and mitochondrial DNA release can activate this pathway, thereby inducing type I interferon response and affecting the composition of tumor-infiltrating immune cells and the function of CD4+ T cells [29].
cAMP signaling also has important significance in the immune regulation of CD4+ T cells. Relevant studies have shown that cAMP pathway activators such as Forskolin can regulate the CD4+ T cell response in autoimmune inflammation, and are associated with the remission of central nervous system inflammation [32]. In addition, there is extensive crosstalk between multiple signaling pathways, which together determine the functional state of CD4+ T cells under different disease backgrounds [28].
HIV infection most typically reflects the core position of CD4 in diseases. CD4 is not only an important indicator for immune function evaluation, but also an important related molecule for HIV to invade host cells. The decrease in the number of CD4+ T cells directly reflects the degree of immune system damage, and factors such as viral load, host individual differences and immune reconstitution ability jointly determine the speed of disease progression. Therefore, CD4 count has long been an important parameter for HIV infection management and efficacy evaluation.
In diseases such as rheumatoid arthritis, systemic lupus erythematosus, and experimental autoimmune encephalomyelitis, the imbalance of CD4+ T cell subsets is one of the key pathological mechanisms. In particular, the imbalance of Th17/Treg is often associated with chronic inflammation amplification and tissue damage [9-10]. A variety of metabolic interventions, cell therapies and signaling pathway regulation strategies have shown potential in alleviating diseases by remodeling CD4+ T cell function [26],[30].
In tumor immunity, CD4+ T cells can assist CD8+ T cells to exert killing effect, and can also independently mediate anti-tumor immunity, and even act as a prognostic predictive biomarker. This indicates that CD4+ T cells are not only "helpers" in tumor immunity, but may also be key "executors" under certain conditions.
In addition to HIV, CD4+ T cells also play an important role in acute and chronic infections such as influenza. Especially the discovery of CD4 CTL suggests that CD4+ T cells are not only responsible for regulation in the antiviral process, but can also directly participate in the clearance of infected cells [1]. This expands the understanding of the functional boundary of CD4+ T cells in infection immunity.
At present, there are various types of drugs targeting CD4, including monoclonal antibodies, bispecific antibodies, CAR-T and small molecule drugs; the researched indications are mainly HIV infection, and expanded to fields such as atopic dermatitis and tumors; the research and development stage covers marketed varieties to early clinical stage, and some of the researched pipelines are listed below.
| Drug | Target | Drug Type | Indications | Research Institutions | Highest Development Stage |
|---|---|---|---|---|---|
| Ibalizumab | CD4 x Viral fusion proteins | Bispecific antibody | HIV infection | Theratechnologies, Inc. | Approved |
| Semzuvolimab | CD4 | Monoclonal antibody | HIV infection | United BioPharma (Great China) Holdings Co., Ltd. | United Biopharma Corporation | United BioPharma, Inc. | Phase 3 |
| Mosedipimod | CD4 x CD8 | Small molecule drug | Moderate atopic dermatitis | Severe atopic dermatitis | Metabolic dysfunction-associated steatohepatitis (MASH) | ENZYCHEM LIFESCIENCES Corp. | Phase 2 |
| Iotivibart | CD4 x HIV envelope protein gp120 | Monoclonal antibody | HIV infection | ViiV Healthcare UK Ltd. | ViiV Healthcare Ltd. | Phase 2 |
| TMB-365 | CD4 | Monoclonal antibody | HIV infection | TaiMed Biologics Inc. | Phase 2 |
| IB-MS | CD4 x CD8 | Small molecule drug | Multiple sclerosis | INNOBIOSCIENCE LLC | Phase 2 |
| CD4 CAR T cell therapy (iCell Gene Therapeutics) | CD4 | Autologous CAR-T | Chronic myelomonocytic leukemia | T-cell leukemia | T-cell lymphoma | iCell Gene Therapeutics, Inc. | Phase 1 |
| IT-1208 (Kyowa Hakko Kirin Pharma, Inc.) | CD4 | Monoclonal antibody | Cancer | IDAC Theranostics, Inc. | Ono Pharmaceutical Co., Ltd. | Kyowa Kirin Co., Ltd. | Phase 1 |
| Autologous CD4 CAR T-cells (Indiana University) | CD4 | Autologous CAR-T | Refractory acute myeloid leukemia | Relapsed acute myeloid leukemia | iCell Gene Therapeutics, Inc. | Phase 1 |
| VRCHIVMAB0115-00-AB | CD4 | Monoclonal antibody | HIV infection | National Institute of Allergy & Infectious Diseases (NIAID) | Phase 1 |
| BG-8962 | CD4 | Biologic | HIV infection | National Institute of Allergy & Infectious Diseases (NIAID) | Phase 1 |
| LM49 | CD4 | Small molecule drug | Diabetic nephropathy | Shanxi Medical University | Shanxi University of Chinese Medicine | Phase 1 |
(Data as of March 16, 2026, sourced from synapse)
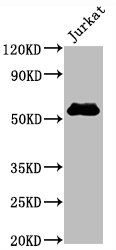
CD4 is not only an important surface molecule on helper T cells, but also a key hub connecting antigen recognition, signal transduction, cell differentiation and disease progression. Research around CD4 and CD4+ T cells has expanded from the traditional "helper function" to multiple levels such as cytotoxicity, metabolic regulation, tissue microenvironment adaptation and disease persistence mechanism. CUSABIO provides CD4 recombinant proteins, antibodies and ELISA kit products to assist you in related mechanism research and targeted drug development.
● CD4 Recombinant Proteins
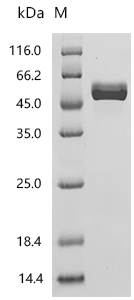
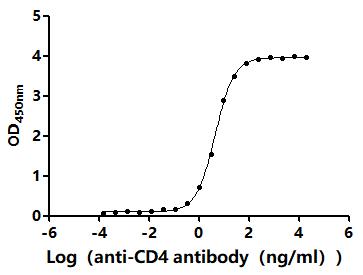
Recombinant Human T-cell surface glycoprotein CD4 (CD4), partial (Active); CSB-MP004935HU1
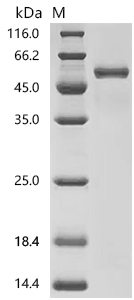
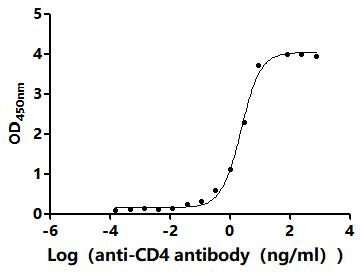
Recombinant Macaca mulatta T-cell surface glycoprotein CD4 (CD4), partial (Active); CSB-MP6740MOW
● CD4 Antibodies
CD4 Recombinant Monoclonal Antibody; CSB-RA004935A0HU
● CD4 ELISA Kits
References
[1] Deborah M. Brown, Anna T. Lampe, Aspen M. Workman. (2016). The Differentiation and Protective Function of Cytolytic CD4 T Cells in Influenza Infection.
[2] Tomas Lukes, Daniela Glatzova, Zuzana Kvicalova, Florian Levet, Ales Benda, Tomas Brdicka, Theo Lasser, Marek Cebecauer. (2017). Quantifying protein densities on cell membranes using super-resolution optical fluctuation imaging.
[3] Mikaël Simard, Zhuoyan Shen, Konstantin Bräutigam, Rasha Abu-Eid, Maria A. Hawkins, Charles-Antoine Collins-Fekete. (2024). Immunocto: a massive immune cell database auto-generated for histopathology.
[4] Xin Shu, Sameera Sansre, Di Jin, Xiangxiang Zeng, Kai-Yu Tong, Rishikesh Pandey, Renjie Zhou. (2020). Artificial Intelligence Enabled Reagent-free Imaging Hematology Analyzer.
[5] D. Rider, C. Havenith, R. De Ridder, J. Schuurman, C. Favre, J. Cooper, S. Walker, O. Baadsgaard, S. Marschner, Jan G J vandeWinkel, J. Cambier, P. Parren, D. Alexander. (2007). A human CD4 monoclonal antibody for the treatment of T-cell lymphoma combines inhibition of T-cell signaling by a dual mechanism with potent Fc-dependent effector activity.
[6] J. T. Ulrich-Lewis, K. Draves, Kelsey Roe, M. O'Connor, E. Clark, D. Fuller. (2022). STING Is Required in Conventional Dendritic Cells for DNA Vaccine Induction of Type I T Helper Cell- Dependent Antibody Responses.
[7] Constance Renault, N. Veyrenche, F. Mennechet, A. Bedin, J. Routy, P. Van de Perre, J. Reynes, E. Tuaillon. (2022). Th17 CD4+ T-Cell as a Preferential Target for HIV Reservoirs.
[8] Rong Wang, Keliang Huang. (2020). CCL11 increases the proportion of CD4+CD25+Foxp3+ Treg cells and the production of IL-2 and TGF-β by CD4+ T cells via the STAT5 signaling pathway.
[9] Jiaxu Qin, Xiangwei Zheng, Yancheng He, Yang Hong, Shuang Liang, X. Fang. (2022). The regulation of T helper cell polarization by the diterpenoid fraction of Rhododendron molle based on the JAK/STAT signaling pathway.
[10] Fanping Wang, Yonghui Yang, Zhixin Li, Yan Wang, Zhenchao Zhang, Wei Zhang, Yonghui Mu, Jing-ke Yang, Lili Yu, Mingyong Wang. (2022). Mannan-Binding Lectin Regulates the Th17/Treg Axis Through JAK/STAT and TGF-β/SMAD Signaling Against Candida albicans Infection.
[11] R. Deng, C. Hurtz, Qingxiao Song, Chanyu Yue, G. Xiao, Hua Yu, Xiwei Wu, M. Muschen, S. Forman, P. Martin, D. Zeng. (2017). Extrafollicular CD4+ T-B interactions are sufficient for inducing autoimmune-like chronic graft-versus-host disease.
[12] Laura M. Fahey, Elizabeth B. Wilson, Heidi J. Elsaesser, Chris Fistonich, D. McGavern, D. Brooks. (2011). Viral persistence redirects CD4 T cell differentiation toward T follicular helper cells.
[13] A. Dejean, Emeline Joulia, T. Walzer. (2018). The role of Eomes in human CD4 T cell differentiation: A question of context.
[14] Yuki Sato, Abhinav Jain, Shozo Ohtsuki, Hirohisa Okuyama, Ines Sturmlechner, Yoshinori Takashima, Kevin-Phu C. Le, Melanie C. Bois, Gerald J. Berry, K. Warrington, J. Goronzy, C. Weyand. (2023). Stem-like CD4+ T cells in perivascular tertiary lymphoid structures sustain autoimmune vasculitis.
[15] S. Keck, M. Schmaler, S. Ganter, Lena Wyss, Susanne G. Oberle, E. Huseby, D. Zehn, C. King. (2014). Antigen affinity and antigen dose exert distinct influences on CD4 T-cell differentiation.
[16] M. Martínez-Sánchez, L. Huerta, E. Álvarez-Buylla, Carlos Villarreal Luján. (2018). Role of Cytokine Combinations on CD4+ T Cell Differentiation, Partial Polarization, and Plasticity: Continuous Network Modeling Approach.
[17] E. Zorn, E. Nelson, M. Mohseni, F. Porcheray, Haesook T. Kim, Despina Litsa, R. Bellucci, E. Raderschall, C. Canning, R. Soiffer, D. Frank, J. Ritz. (2006). IL-2 regulates FOXP3 expression in human CD4+CD25+ regulatory T cells through a STAT-dependent mechanism and induces the expansion of these cells in vivo.
[18] M. Kroenke, D. Eto, M. Locci, Michael Cho, T. Davidson, E. Haddad, S. Crotty. (2012). Bcl6 and Maf Cooperate To Instruct Human Follicular Helper CD4 T Cell Differentiation.
[19] Megan A. Luckey, M. Kimura, A. Waickman, L. Feigenbaum, A. Singer, Jung-Hyun Park. (2014). The transcription factor ThPOK suppresses Runx3 and imposes CD4+ lineage fate by inducing the SOCS suppressors of cytokine signaling.
[20] Qingtian Li, Jia Zou, Mingjun Wang, Xilai Ding, I. Chepelev, Xikun Zhou, Wei Zhao, Gang Wei, J. Cui, K. Zhao, Helen Y Wang, Rong-Fu Wang. (2014). Critical role of histone demethylase Jmjd3 in the regulation of CD4+ T-cell differentiation.
[21] Y. Shu, Qinghua Hu, H. Long, Christopher Chang, Q. Lu, R. Xiao. (2017). Epigenetic Variability of CD4+CD25+ Tregs Contributes to the Pathogenesis of Autoimmune Diseases.
[22] Sarah A. LaMere, Ryan C. Thompson, Xiangzhi Meng, H. Komori, Adam M. Mark, D. Salomon. (2017). H3K27 Methylation Dynamics during CD4 T Cell Activation: Regulation of JAK/STAT and IL12RB2 Expression by JMJD3.
[23] M. Martínez-Sánchez, L. Mendoza, C. Villarreal, E. Álvarez-Buylla. (2015). A Minimal Regulatory Network of Extrinsic and Intrinsic Factors Recovers Observed Patterns of CD4+ T Cell Differentiation and Plasticity.
[24] B. L. Puniya, R. G. Todd, Akram Mohammed, Deborah M. Brown, Matteo Barberis, T. Helikar. (2018). A Mechanistic Computational Model Reveals That Plasticity of CD4+ T Cell Differentiation Is a Function of Cytokine Composition and Dosage.
[25] V. Proserpio, A. Piccolo, Liora Haim-Vilmovsky, G. Kar, T. Lönnberg, Valentine Svensson, J. Pramanik, K. Natarajan, Weichao Zhai, Xiuwei Zhang, G. Donati, M. Kayikci, J. Kotar, A. McKenzie, Ruddy Montandon, O. Billker, S. Woodhouse, P. Cicuta, M. Nicodemi, S. Teichmann. (2016). Single-cell analysis of CD4+ T-cell differentiation reveals three major cell states and progressive acceleration of proliferation.
[26] S. Tullius, H. Biefer, Suyan Li, A. Trachtenberg, K. Edtinger, M. Quante, Felix Krenzien, H. Uehara, Xiaoyong Yang, H. Kissick, W. Kuo, I. Ghiran, M. A. de la Fuente, M. Arredouani, Virginia Camacho, J. Tigges, Vasilis Toxavidis, Rachid El Fatimy, Brian D. Smith, A. Vasudevan, A. Elkhal. (2014). NAD+ protects against EAE by regulating CD4+ T-cell differentiation.
[27] Nizar Chetoui, M. Boisvert, S. Gendron, F. Aoudjit. (2010). Interleukin‐7 promotes the survival of human CD4+ effector/memory T cells by up‐regulating Bcl‐2 proteins and activating the JAK/STAT signalling pathway.
[28] M. S. Alam, Y. Maekawa, A. Kitamura, K. Tanigaki, T. Yoshimoto, K. Kishihara, K. Yasutomo. (2010). Notch signaling drives IL-22 secretion in CD4+ T cells by stimulating the aryl hydrocarbon receptor.
[29] Shengpan Wu, Baojun Wang, Hong-zhao Li, Hanfeng Wang, Songliang Du, Xing Huang, Yang Fan, Yu Gao, L. Gu, Qingbo Huang, Jianjun Chen, Xu Zhang, Yan Huang, Xin Ma. (2024). Targeting STing elicits GSDMD-dependent pyroptosis and boosts anti-tumor immunity in renal cell carcinoma.
[30] Dongmei Li, Idalia Cruz, Yahui Feng, Maha Moussa, Jie Cheng, Digvijay Patil, Alexander Kroemer, Joseph A. Bellanti. (2025). Divergent Immunomodulatory Roles of Fungal DNA in Shaping Treg and Inflammatory Responses.
[31] C. Elsner, Aparna Ponnurangam, J. Kazmierski, T. Zillinger, Jenny Jansen, D. Todt, K. Döhner, Shuting Xu, A. Ducroux, Nils Kriedemann, Angelina Malassa, P. Larsen, G. Hartmann, W. Barchet, E. Steinmann, U. Kalinke, B. Sodeik, C. Goffinet. (2020). Absence of cGAS-mediated type I IFN responses in HIV-1--infected T cells.
[32] Tatyana Veremeyko, Amanda W. Y. Yung, M. Dukhinova, Inna S. Kuznetsova, I. Pomytkin, A. Lyundup, T. Strekalova, N. Barteneva, E. Ponomarev. (2018). Cyclic AMP Pathway Suppress Autoimmune Neuroinflammation by Inhibiting Functions of Encephalitogenic CD4 T Cells and Enhancing M2 Macrophage Polarization at the Site of Inflammation.


Comments
Leave a Comment