Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
Fibroblast growth factor 2 (FGF2) is a multifunctional growth factor widely involved in processes such as cell proliferation, angiogenesis, tissue repair, and tumor progression. This article systematically reviews the molecular structure, signaling pathways (e.g., PI3K/AKT/mTOR, MAPK/ERK), and the mechanisms of FGF2 action in diseases including cancer, neurological disorders, retinopathies, and skeletal conditions, aiming to provide a reference for mechanism-based research and precision therapy in FGF2-related diseases.
1. Introduction: Overview and Biological Significance of FGF2
2. Molecular Biological Characteristics and Mechanism of Action of FGF2
3. FGF2 Signaling Transduction Pathways and Regulatory Networks
4. Pathophysiology of FGF2-Related Diseases
Fibroblast growth factor 2 (FGF2), also known as basic fibroblast growth factor (bFGF), is an important member of the FGF family that plays a key role in cell proliferation, differentiation, migration, angiogenesis and tissue repair. The FGF family consists of structurally conserved peptide growth factors that primarily bind to fibroblast growth factor receptors (FGFRs) and heparan sulfate (HS) proteoglycans, activating downstream signaling pathways and regulating cell behavior [1, 2]. Studies have shown that FGF2 can promote FGFR conformational changes and enhance its activation level, and HS and its modifications also play an important role in FGF2/FGFR signaling regulation [1-3].
FGF2 is involved in multiple key biological processes. In terms of cell proliferation, YAP can enhance the response of neural stem cells to FGF2 by upregulating FGFR expression; in ER-positive breast cancer, FGF2 can also activate ERα and PR signaling, promoting tumor cell proliferation and inducing endocrine therapy resistance [4, 5]. In angiogenesis, FGF2 is an important pro-angiogenic factor that often cooperates with molecules such as VEGF to participate in vessel formation and remodeling, and plays a role in myocardial injury repair, retinopathy, and ischemic stroke [6-10]. In differentiation regulation, FGF2 is closely associated with osteogenesis and odontogenic differentiation, while also exhibiting positive effects in tissue repair and anti-fibrosis [11-15].
FGF2 also exists in different isoforms such as low molecular weight (LMW) and high molecular weight (HMW), which differ in intracellular localization and function, and may play different roles in disease processes [16]. Meanwhile, abnormal expression or signaling imbalance of FGF2 is closely related to various diseases, particularly involving tumor progression, therapeutic resistance, and some inflammation-related diseases [17-21]. Overall, FGF2 has both important basic research value and clinical translational potential. In-depth study of its mechanism of action, signaling pathways, and disease associations is of great significance for understanding the development of related diseases and exploring new therapeutic strategies.
FGF2 is a functionally diverse and important member of the FGF family, and its protein structure shares the common family feature of a highly conserved central β-barrel structure, which is crucial for its binding to FGFR and heparan sulfate-related molecules. The biological complexity of FGF2 largely arises from its multiple protein isoforms. The Fgf2 gene can be translated through different initiation codons to generate low molecular weight (approximately 18 kDa) and high molecular weight (approximately 22--24 kDa) isoforms [16, 23]. Among these, LMW FGF2 is mainly localized in the cytoplasm and nucleus, while HMW FGF2 is more concentrated in the nucleus. The differences in intracellular localization and function between different isoforms allow FGF2 to exhibit more complex regulatory patterns in different tissues and disease contexts.
It is worth noting that FGF2 lacks a classical signal peptide, so its secretion does not depend on the traditional endoplasmic reticulum-Golgi pathway, but is released to the extracellular space through non-classical secretion. This characteristic determines that FGF2 has unique properties in terms of deposition in the extracellular matrix, local concentration formation, and paracrine/autocrine regulation, which provides a basis for its spatiotemporally specific role in development, repair, and disease.
FGF2 signal transduction begins with its specific binding to cell surface FGFRs, a process that typically relies on the cooperation of heparan sulfate proteoglycans (HSPGs). The formation of a functional ternary complex by FGF2, FGFR, and HSPG is considered the basis for FGF2 biological activity. After FGF2 binds to the receptor, it promotes FGFR dimerization and triggers phosphorylation of the intracellular tyrosine kinase domain, subsequently initiating a series of downstream signaling cascades.
However, the interaction between FGF2 and HS/HSPGs is not simply a "presence or absence" relationship. Previous studies have shown that reducing the binding ability of FGF2 to HS does not completely eliminate FGFR1 signaling [2], indicating that the contribution of co-receptors may be context-dependent. Meanwhile, specific types of HS modifications, such as 2-O sulfation, are still crucial for effective activation of the FGF2/FGFR1/Erk1/2 pathway [3]. Therefore, the formation and activation of the FGF2 receptor complex is not a single molecular event, but a finely regulated process jointly controlled by ligands, receptors, and the glycosaminoglycan microenvironment.
FGF2 is involved in regulating multiple key cellular behaviors, including cell proliferation, migration, differentiation, anti-apoptosis, and angiogenesis. In stem cell maintenance and proliferation, FGF2 is important for maintaining the stemness of cell populations such as bone marrow mesenchymal stem cells; in the nervous system, YAP promotes neural stem cell expansion by enhancing cellular response to FGF2 [4]. In the tumor microenvironment, FGF2 can drive tumor cell proliferation and promote the formation of therapeutic resistance [5, 17].
In terms of differentiation, the regulation of FGF2 on osteogenesis and odontogenic differentiation is particularly prominent. It can enhance osteogenic differentiation through the ERK-Runx2 axis [13], and promote osteogenic/odontogenic differentiation of SCAP by inhibiting the PI3K/AKT pathway in specific cellular contexts [11]. In tissue repair and angiogenesis, FGF2 can cooperate with factors such as VEGF to enhance neovascularization, improve local blood supply, and promote the repair of damaged tissues [6, 8, 10, 14]. Therefore, FGF2 is not merely a single-function growth factor, but an important regulatory node throughout multiple cellular processes.
FGF2 activates multiple intracellular signaling pathways through binding to FGFR, among which the PI3K/AKT/mTOR pathway is one of its core downstream pathways. After FGFR activation, it recruits and activates PI3K, promoting the conversion of PIP2 to PIP3, thereby further activating AKT. Activated AKT can regulate mTOR and other downstream effector molecules, ultimately affecting cell growth, metabolism, survival, anti-apoptosis and other processes.
In different disease and tissue contexts, the regulation of the PI3K/AKT/mTOR pathway by FGF2 is diverse. For example, in pathological angiogenesis, downregulation of PIK3IP1 can enhance PI3K-AKT signaling and increase the expression of pro-angiogenic factors such as FGF2, thereby exacerbating abnormal angiogenesis [9]. In tissue repair, the CXCR4/Akt/FGF2 axis can promote FGF2 secretion and enhance tissue regeneration [14]. However, in certain differentiation scenarios, FGF2 may promote differentiation by inhibiting the PI3K/AKT pathway, as seen in osteogenic/odontogenic differentiation of SCAP [11]. This indicates that the relationship between FGF2 and the PI3K/AKT/mTOR pathway is not unidirectional activation, but has obvious cell type and biological context dependence.
The MAPK/ERK pathway is another classic pathway mediating FGF2 cellular effects. After FGF2 binds to FGFR, it activates ERK through adaptor proteins and the Ras-Raf-MEK cascade reaction, thereby regulating cell growth, differentiation, migration, and stress responses. YAP can enhance cellular response to FGF2 by upregulating FGFR1-4 expression, and further activate MAPK and AKT signaling, thereby promoting embryonic neural stem cell proliferation [4]. During bone formation, FGF2 activates ERK to enhance the protein stability and transcriptional activity of Runx2, playing a positive role in osteoblast differentiation and bone formation [13].
However, the MAPK/ERK pathway is also one of the important mechanisms for FGF2 to participate in tumor drug resistance. In KIT-mutant melanoma, the FGF2 axis promotes resistance to nilotinib by inhibiting BIM expression through the MAPK pathway [18]. In glioma, radiation-induced YAP activation can upregulate FGF2 and further enhance MAPK-ERK signaling, forming radioresistance [19]. Therefore, the FGF2-MAPK/ERK axis is both an important pathway for normal physiological regulation and an important molecular basis for tumor progression and treatment failure.
The FGF2 signaling network does not operate independently, but undergoes complex cross-regulation with multiple signaling pathways such as VEGF/VEGFR, Src, YAP, and CXCR4. Taking angiogenesis as an example, FGF2 and VEGF often jointly participate in vessel formation and remodeling, showing obvious synergistic effects in ischemic repair and retinopathy [6, 7, 8, 10]. In pathological environments, Src pathway activation is accompanied by increased expression of FGF2 and its receptors, enhancing the pro-angiogenic effect [8]. In addition, YAP can both enhance cellular response to FGF2 by upregulating FGFR and promote FGF2 secretion after radiotherapy, forming a pro-survival and drug resistance loop [4, 19].
This cross-regulation further illustrates that FGF2 is not a linear factor in a single pathway, but a key hub in a multi-layered regulatory network. It is precisely because of this that therapeutic strategies targeting FGF2 are often difficult to achieve ideal results through a single node, and combined blockade of multiple nodes often has greater potential.
FGF2 plays an important pro-tumor role in the development of various tumors, with mechanisms involving cell proliferation, migration, invasion, angiogenesis, and multiple types of therapeutic resistance. In ER-positive breast cancer, FGF2 can promote tumor growth by activating estrogen-independent ER signaling and lead to endocrine therapy resistance [5, 17]. In KIT-mutant melanoma, the FGF2 axis is involved in the formation of nilotinib resistance, which can be partially reversed by MEK inhibitors [18]. In non-small cell lung cancer, breast cancer, and melanoma, high expression of FGF2/FGFR1 is associated with DTP formation after TKI treatment, suggesting that compensatory activation of FGFR signaling is one of the important mechanisms for acquired drug resistance in tumors.
In addition to drug resistance, FGF2 is also closely associated with tumor invasiveness. Its systemic level is elevated in some patients with gastric and esophageal cancer, which may reflect a stronger tendency for disease progression [20]. In glioma, FGF2 is involved in the formation of radioresistance, making it a potential radiosensitization target [19]. Therefore, FGF2 is not only a tumor growth-related factor, but also an important molecule affecting treatment response and clinical outcome.
FGF2 has multiple roles in the nervous system, including promoting proliferation, nutritional support, and tissue protection. Studies have shown that YAP can promote the proliferation of neural stem cells by enhancing their response to FGF2 [4]. In addition, FGF2 exhibits neurotrophic and neuroprotective effects in Parkinson's disease models, supporting the survival of dopaminergic neurons and reducing damage induced by MPP+ or rotenone.
In retinopathy, FGF2 is involved in tissue repair and is also closely related to pathological angiogenesis. Under hypoxic conditions, YY1 lactylation can enhance FGF2 transcription and promote angiogenesis [7]; downregulation of PIK3IP1 and deletion of the miRNA-15a/16-1 cluster can both enhance FGF2-related signaling through different mechanisms, driving abnormal angiogenesis [8, 9]. Meanwhile, A1-PEG can improve vascular repair and retinal function by increasing FGF2 expression while inhibiting pathological neovascularization [10]. This indicates that FGF2 has obvious duality in neurological and retinal diseases, and its direction of action is highly dependent on the specific pathological context.
FGF2 plays an important role in bone formation, remodeling, and maintenance. It enhances the stability and transcriptional activity of Runx2 by activating ERK MAP kinase, thereby promoting osteoblast differentiation and bone morphogenesis [13]. In tooth-related tissues, FGF2 can also enhance the osteogenic/odontogenic differentiation capacity of SCAP, which provides an important basis for pulp-dentin repair research [11].
On the other hand, FGF2 signaling imbalance is also associated with abnormal bone metabolism. For example, sodium benzoate can inhibit the FGF2/p38/RUNX2 pathway, leading to impaired osteogenesis and bone loss [12]. In addition, FGF2 is also important in connective tissue repair and anti-fibrosis, and its derived short peptide P5 can reduce EMT and collagen deposition in a pulmonary fibrosis model [15]. Therefore, FGF2 is both an important regulator of pathological changes and a potential intervention target in bone, dental, and connective tissue diseases.
The role of FGF2 in inflammatory, infectious, cardiovascular, and metabolic diseases is also complex. In cardiovascular injury repair, FGF2, together with factors such as VEGF, participates in angiogenesis, improving blood supply to the infarcted myocardium and promoting functional recovery [6]. In radiation-induced intestinal injury, the CXCR4/Akt/FGF2 axis helps promote intestinal regeneration [14]. In a myocardial infarction model, different FGF2 isoforms have different effects on long-term cardiac function and angiogenesis, suggesting that isoform differences may affect disease outcomes [16].
In inflammatory and infectious diseases, the role of FGF2 is gradually receiving attention. Proteomic studies have suggested a causal association between FGF2 and periodontitis [21]. In metabolically related pathological angiogenesis, such as early diabetic retinopathy, FGF2 and VEGF jointly participate in disease progression. Overall, the role of FGF2 in these diseases is not simply beneficial or harmful, but has obvious environment and stage dependence.
Globally, R&D on drugs targeting bFGF (FGF2) and related pathways is diversifying, primarily along two fronts: promoting regeneration (using gene therapies and novel delivery systems for chronic wounds and ischemia) and inhibiting tumors (developing small molecules and biologics targeting FGF/FGFR signaling for solid tumors). The field is evolving toward highly selective inhibitors and novel technologies like PROTACs to overcome resistance. Selected pipelines are listed below:
| Drug Name | Target | Drug Type | Indications (Disease Name) | Institutions | Highest Development Phase |
|---|---|---|---|---|---|
| Iremythelin Alfa | VEGF x bFGF | Fc Fusion Protein | Diabetic Macular Edema; Wet Age-Related Macular Degeneration | Rongchang Biopharmaceutical (Yantai) Co., Ltd. | Application for Marketing |
| Sulfonated Manno-Oligosaccharide | VEGF-A x aFGF x bFGF | Polymer | Hepatocellular Carcinoma | Kylin Biotech Co., Ltd. | Phase 3 |
| AUP1602-C | M-CSF x bFGF | Engineered Bacteria | Diabetic Foot Ulcer; Diabetic Foot; Venous Leg Ulcer; Pressure Ulcer | Shenzhen UnknownKey Biotechnology Co., Ltd.; Aurealis Oy; Aurealis Therapeutics AG | Phase 2 |
| Curcumin C3 Complex | NF-κB x bFGF x mTOR | Small Molecule Drug | Cervical Intraepithelial Neoplasia; Human Papillomavirus-Related Cervical Cancer; Cutaneous Squamous Cell Carcinoma | Emory University; LSU Health Shreveport | Phase 2 |
| Umedaptanib Pegol | bFGF | Aptamer | Achondroplasia; Diabetic Macular Edema | Ribomic, Inc. | Phase 2 |
| Oleandrin | AP-1 x FLI1 x Na/K-ATPase x RPS6KB1 x bFGF x mTOR | Small Molecule Drug | Pancreatic Cancer; Neoplasms | U.S. Army Medical Research Institute for Infectious Diseases; Nerium Biotechnology, Inc.; The University of Texas MD Anderson Cancer Center | Phase 2 |
| Chlorogenic Acid | Akt x DNMT1 x LAG3 x SLC37A4 x TGF-β1 x bFGF | Small Molecule Drug | Recurrent Glioma; Advanced Lung Adenocarcinoma; Advanced Small Cell Lung Cancer; Advanced Squamous Cell Lung Cancer; Renal Tumors, etc. | Sichuan Jiuzhang Biotech Co., Ltd.; Hunan University of Chinese Medicine; Universiti Putra Malaysia; Brawijaya University | Phase 2 |
| Fibroblast Growth Factor 2 Gene Therapy (Id Pharma Co., Ltd.) | bFGF | Gene Therapy | Intermittent Claudication; Peripheral Arterial Disease; Perioperative Ischemia | Kyushu University; Id Pharma Co., Ltd. | Phase 2 |
| rHIgM22 | PDGFRs x bFGF | Monoclonal Antibody | Demyelinating Diseases | Mayo Clinic | Phase 1 |
(Data as of March 19, 2026, from Synapse)
FGF2 is a pleiotropic growth factor with extensive biological functions that plays a central role in cell proliferation, differentiation, migration, angiogenesis, and tissue repair. By binding to FGFRs, it activates MAPK/ERK, PI3K/AKT, and multiple other signaling pathways, forming a complex and finely regulated network. Meanwhile, abnormal expression or signaling dysregulation of FGF2 is closely associated with various diseases, particularly prominent in tumor progression, therapeutic resistance, retinopathy, neurological disorders, abnormal bone metabolism, and fibrosis. CUSABIO offers FGF2 recombinant proteins, antibodies, and ELISA kits to support your research on related mechanisms and targeted drug development.
● FGF2 Recombinant Protein
Recombinant Bovine Fibroblast growth factor 2 (FGF2) (Active); CSB-EP008625BOe1
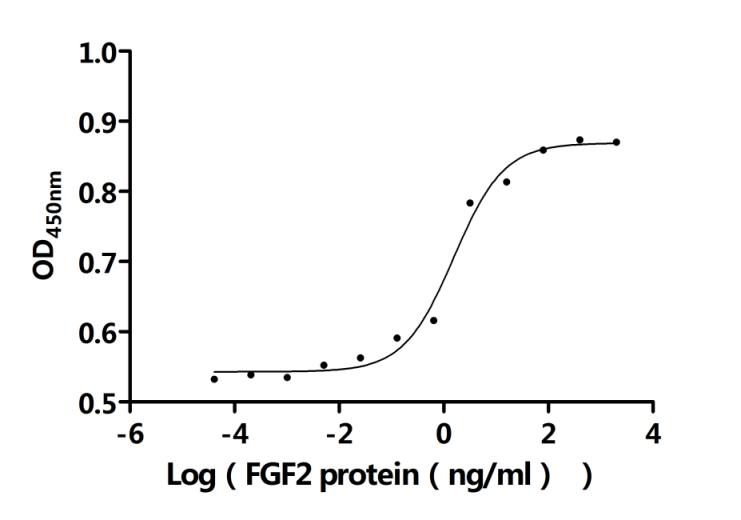
● FGF2 ELISA Kits
References
[1] S. Sarabipour, K. Hristova. (2016). Mechanism of FGF receptor dimerization and activation.
[2] Yuga Okada, Akihiro Eguchi, Daisuke Kuroda, Kouhei Tsumoto, Ryosuke Ueki, Shinsuke Sando. (2025). Developing FGF2 Mutants with Selectively Reduced Heparan Sulfate Affinity to Explore Their Impact on FGFR1 Signaling.
[3] W. Chan, K. Howe, James M. Clegg, S. Guimond, D. Price, J. Turnbull, T. Pratt. (2015). 2-O Heparan Sulfate Sulfation by Hs2st Is Required for Erk/Mapk Signalling Activation at the Mid-Gestational Mouse Telencephalic Midline.
[4] Dasol Han, S. Lee, Mookwang Kwon, Hogyun Noh, J. H. Lee, Y. Yoon, J. Cho, K. Yoon. (2020). YAP enhances FGF2-dependent neural stem cell proliferation by induction of fibroblast growth factor receptor expression.
[5] Sebastián Giulianelli, Marina Riggio, Tomás Guillardoy, C. Pérez Piñero, M. Gorostiaga, G. Sequeira, Gabriela Pataccini, M. F. Abascal, Maria F. Toledo, B. Jacobsen, Ana C. L. Guerreiro, António S. Barros, V. Novaro, F. Monteiro, Francisco Amado, H. Gass, M. Abba, L. Helguero, C. Lanari. (2019). FGF2 induces breast cancer growth through ligand‑independent activation and recruitment of ERα and PRBΔ4 isoform to MYC regulatory sequences.
[6] Bin Zeng, Guosheng Lin, X. Ren, Yan Zhang, Honglei Chen. (2010). Over-expression of HO-1 on mesenchymal stem cells promotes angiogenesis and improves myocardial function in infarcted myocardium.
[7] Xiaotang Wang, W. Fan, Na Li, Yan Ma, Mudi Yao, Guoqing Wang, Siyuan He, Wanqian Li, J. Tan, Qingjun Lu, Shengping Hou. (2021). YY1 lactylation in microglia promotes angiogenesis through transcription activation-mediated upregulation of FGF2.
[8] P. Sun, Feifei Ma, Yang Xu, Chao Zhou, R. A. Stetler, K. Yin. (2021). Genetic deletion of endothelial microRNA-15a/16-1 promotes cerebral angiogenesis and neurological recovery in ischemic stroke through Src signaling pathway.
[9] Lushu Chen, Yuan Cao, Yaming Shen, Huan Li, Rong Ye, Jin Yao. (2023). Downregulation of PIK3IP1 in retinal microglia promotes retinal pathological neovascularization via PI3K-AKT pathway activation.
[10] A. Fouda, Zhimin Xu, J. Suwanpradid, M. Rojas, E. Shosha, Tahira Lemtalsi, C. Patel, Ji Xing, Sumbal Zaidi, W. Zhi, Brain K Stansfield, P. Cheng, S. Narayanan, R. Caldwell, R. Caldwell. (2022). Targeting proliferative retinopathy: Arginase 1 limits vitreoretinal neovascularization and promotes angiogenic repair.
[11] Zijie Wang, Chuying Chen, Liying Sun, Mei He, Ting Huang, Jiji Zheng, Jiayuan Wu. (2024). Fibroblast growth factor 2 promotes osteo/odontogenic differentiation in stem cells from the apical papilla by inhibiting PI3K/AKT pathway.
[12] Zhonghao Wang, Yexin Wang, Yu Tang, Xiaoyan Guo, Qize Gao, Yiming Shao, Jingxuan Wang, Ronghua Tian, Yingxu Shi. (2025). Sodium Benzoate Inhibits Osteoblast Differentiation and Accelerates Bone Loss by Regulating the FGF2/p38/RUNX2 Pathway.
[13] Ok-Jin Park, Hyun-Jung Kim, K. Woo, J. Baek, H. Ryoo. (2009). FGF2-activated ERK Mitogen-activated Protein Kinase Enhances Runx2 Acetylation and Stabilization.
[14] Yang Jiao, Jing Xu, Bingtan Song, Ailing Wu, L. Pan, Ying Xu, Fenghao Geng, Xiaoqian Li, Congzhao Zhao, Min Hong, Xuanyu Meng, Judong Luo, Peng Liu, Ming Li, Wei Zhu, Jianping Cao, Shuyu Zhang. (2022). Interferon regulatory factor 1‐triggered free ubiquitin protects the intestines against radiation‐induced injury via CXCR4/FGF2 signaling.
[15] Mengwei Wang, Yuanmeng Sun, Yanzhi Zhao, Xinyi Jiang, Teng Wang, Junye Xie, Xiuling Yu, Shujun Guo, Yibo Zhang, Xiaojia Chen, An Hong. (2025). An FGF2-Derived Short Peptide Attenuates Bleomycin-Induced Pulmonary Fibrosis by Inhibiting Collagen Deposition and Epithelial--Mesenchymal Transition via the FGFR/MAPK Signaling Pathway.
[16] R. Harris, A. Adeyemo, J. Redfield, Cha-hua Huang, Min Jiang, Xiaoping Ren, W. Jones, Jack Rubinstein, J. J. Schultz. (2016). Long‐term Biological Functions of Fibroblast Growth Factor‐2 (FGF2) in Mouse Hearts Following Myocardial Infarction.
[17] Josh W DiGiacomo, Inês Godet, Michael Trautmann-Rodriguez, Daniele M. Gilkes. (2020). Extracellular Matrix--Bound FGF2 Mediates Estrogen Receptor Signaling and Therapeutic Response in Breast Cancer.
[18] P. Têtu, J. Delyon, J. André, Coralie Reger de Moura, Malak Sabbah, G. Ghanem, M. Battistella, S. Mourah, C. Lebbé, N. Dumaz. (2020). FGF2 Induces Resistance to Nilotinib through MAPK Pathway Activation in KIT Mutated Melanoma.
[19] Yu Zhang, Yan Wang, Di Zhou, Kai Wang, Xu Wang, Xiang Wang, Yang Jiang, Min Zhao, Rutong Yu, Xiuping Zhou. (2021). Radiation-induced YAP activation confers glioma radioresistance via promoting FGF2 transcription and DNA damage repair.
[20] Iwona Bednarz-Misa, Paulina Fortuna, D. Diakowska, Natalia Jamrozik, M. Krzystek-Korpacka. (2020). Distinct Local and Systemic Molecular Signatures in the Esophageal and Gastric Cancers: Possible Therapy Targets and Biomarkers for Gastric Cancer.
[21] Z. Alayash, S. Baumeister, B. Holtfreter, Thomas Kocher, H. Baurecht, B. Ehmke, D. Hagenfeld, S. L. Reckelkamm, M. Nolde. (2025). Proteome-Guided Drug Target Discovery for Periodontitis.


Comments
Leave a Comment