Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
| Code | CSB-AP001721HU |
| Abbreviation | Recombinant Human IL5 protein (Active) |
| MSDS | |
| Size | $354 |
| Order now | |
| Image |
|
| Have Questions? | Leave a Message or Start an on-line Chat |
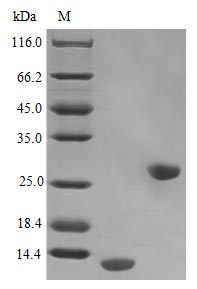
IL-5 is a central driver of eosinophil differentiation, activation, and survival, making it indispensable for studying type 2 immune responses and eosinophilic inflammation. This tag-free recombinant human IL-5, encompassing the full mature sequence (residues 20–134), demonstrates exceptional signaling potency with an ED50 below 0.1 ng/ml in TF-1 cell proliferation assays, corresponding to a specific activity exceeding 1.0 × 10⁷ IU/mg — a level that supports use in eosinophil differentiation and survival assays, JAK/STAT signaling pathway studies, and as a reference standard in cytokine detection platforms. Endotoxin levels below 1.0 EU/μg minimize the risk of LPS-driven artifacts in sensitive immune cell functional assays, including PBMC activation experiments and in vivo models of allergic airway inflammation. Purity exceeding 98% by SDS-PAGE, combined with this stringent endotoxin control, provides a suitable basis for antibody development workflows such as immunogen preparation, ELISA coating, and positive control generation.
There are currently no reviews for this product.

