Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
Lymphocytes are the smallest white blood cells. It is an immune-recognition cell line produced by lymphoid organs. According to their migration, surface makers and biological functions, they can be divided into three categories: T lymphocytes (T cells), B lymphocytes (B cells), and natural killer (NK) cells. These three subpopulations play different roles in immunity: T lymphocytes elicits cell-mediated immunity, B lymphocytes are primarily responsible for humoral immunity, and NK cell mediates innate immunity.
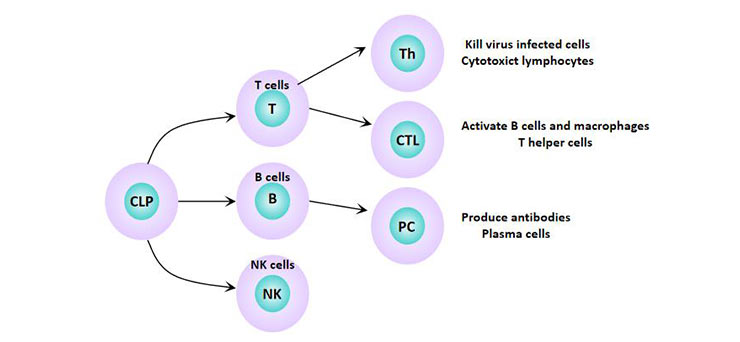
Figure 1 Lymphocyte subsets
1. Why is it necessary to identify different lymphocyte subsets?
2. How Do We identify different lymphocyte subsets?
3. What are Cell Surface Markers?
4. T Lymphocyte and Its Cell Surface Markers
5. B Lymphocyte and Its Cell Surface Markers
The balance of lymphocyte subsets maintains the normal immune function of the body. Abnormal changes in the number and function of lymphocyte subsets will lead to a series of pathological changes and immune dysfunction in the body, resulting in the occurrence of diseases. Studies have shown that lymphocyte subsets play important roles in neoplasia [1] [2] , infectious diseases [3] [4], organ transplantation [5] [6], autoimmune diseases, diabetes, and other processes. Therefore, the identification and monitoring of lymphocyte subsets are of great importance in the clinical.
The three types of lymphocytes are basically similar in morphology and need to distinguish different subpopulations by surface marker detection. Lymphocyte surface markers vary according to lymphocyte type and differentiation stage.
Cell surface markers refer to membrane proteins embedded in the lipid bilayer structure of cell membrane, generally including membrane antigens, membrane receptors, and other molecules. Interactions between lymphocytes and other cells and molecules in the surrounding environment, as well as the biological effects of lymphocytes in antigens, including recognizing, activating, assisting, inhibiting and killing, are all related to their surface markers. The use of cell surface markers to distinguish T lymphocytes from B lymphocytes has greatly promoted the analysis of the cellular basis of immunological phenomena [7].
Detection of lymphocyte surface markers has been widely used in basic and clinical immunological studies and in the determination of patients immune function. Lymphocyte surface markers are generally classified into three major categories: surface immunoglobulin (SIg), differentiation antigen (or specific antigen), and membrane receptors. CD (cluster of differentiation) molecule is one of the most commonly used cell surface markers.
T cells are antigen-specific lymphocytes derived from hematopoietic tissue and mature in the thymus. This is where their name comes from - thymus dependent lymphocytes. T cells can be divided into helper T cells (Th), cytotoxic T cells (Tc), and regulatory T cells (Treg). Changes in the Th/Tc ratio can alter the immune function of the body [8]. When stimulated by antigen, T lymphocytes are transformed into lymphoblasts and then differentiated into sensitized T lymphocytes, which are involved in cellular immunity. It mainly defenses against intracellular infection, tumor cells, and allogeneic cells.
NKT lymphocytes are T lymphocyte subsets with NK cell characteristics [9], which have both the anti-tumor immune activity of T lymphocytes and the non-MHC limiting tumor-killing features of NK cells [10]. And they are important cells in the body against infection, tumor resistance, and regulation of autoimmune balance [11].
In T lymphocytes, helper T cells can be further divided into Th0, Th1, Th2, Th3 (TGF-β), Th17 (IL17), etc.
The distinction between Th1 and Th2 can be made using a relatively specific marker CD antigen:
Th2---CD3+CD4+CD30+
Studies have shown that Th1 has an obvious cytotoxic effect and mainly secretes cytokines such as IL2, IFN-γ, or TNF-β to assist cellular immunity or participate in delayed-type hypersensitivity reaction. Th1 cell surface markers also include IL-12β2R [12] and IL-18R [13]. Th2 does not have cytotoxic effect and primarily secretes cytokines such as IL4, IL5, IL-6 or IL10 to assist humoral immunity and involves in rapid type hypersensitivity.
The study found [14] that Th1/Th2 imbalance disorders are involved in a variety of disease processes, such as tumor immunity, transplant immunity and allergies.
In T lymphocytes, cytotoxic T cells (Tc) can be further divided into:
Tc1 (IFN-γ) and Tc2 (IL-4, IL5, IL-10).
The relatively specific CD antigen markers that distinguish Tc1 from Tc2:
Tc1---CD3+CD8+CD30-
Tc2---CD3+CD8+CD30+
The following table is a surface marker for T lymphocytes.
Table 1 Main CD antigens and their distribution on T cell surface
| CD antigen | Distribution |
|---|---|
| CD2(ER) | All T cells and all NK cells |
| CD3 | Mature T cell |
| CD4(HIVR) | Th cell |
| CD8 | Tc cells, partial NK cells |
| CD25(IL-2R) | Activated T cell |
| CD28 | T cell |
| CTLA-4 | Activated T cell |
| CD40L | Activated T cell |
CD2, also known as LFA2, is an adhesion molecule that binds to sheep red blood cells (SRBC) and is also known as the sheep red blood cell receptor (E receptor). Its ligand is LAF3. It enhances adhesion between T cells and APCs or target cells, and promotes T cell recognition of antigens. Its cytoplasmic domain can be linked to a variety of tyrosine kinases and mediate signaling.
CD3 binds to the TCR transmembrane region to form a stable TCR-CD3 complex. In this complex, the TCR is responsible for recognizing the antigenic peptide-MHC molecular complex, and the CD3 molecule is responsible for transmitting the activation signal into the cell.
The cytoplasmic domain of the CD3 molecule contains an immunoreceptor tyrosine activation motif (ITAM) related to immune cell activation signal transduction, indicating that it is involved in immune cell activation.
CD4 is a T helper cell marker, which is a single chain transmembrane protein. The extracellular structure belongs to IgSF, and there are four IgSF domains. The first and second domains can bind to MHC class II molecules. CD4 acts as a co-receptor for the TCR-CD3 complex recognition antigen and participates in signal transduction by binding to the MHC class II molecule, p56lek kinase.
CD8 is a cytotoxic T cell marker, a heterodimer formed by the linkage of α and β chains by disulfide bonds, and the extracellular structure is an IgSF member. The cytoplasmic region of CD8 molecule can be combined with p56lek kinase to participate in signal transduction. CD8+T lymphocytes, which can specifically kill target cells, have anti-tumor, antiviral and important immunomodulatory effects, and their main function is to inhibit the immune response [15].
CD4 and CD8 molecules divide T cells into two distinct subpopulations. CD4 and CD8 are receptors of MHC class II or MHC class I molecules, respectively, and the changes in the number and ratio of CD4+ and CD8+ cells reflect the immune function status of the body.
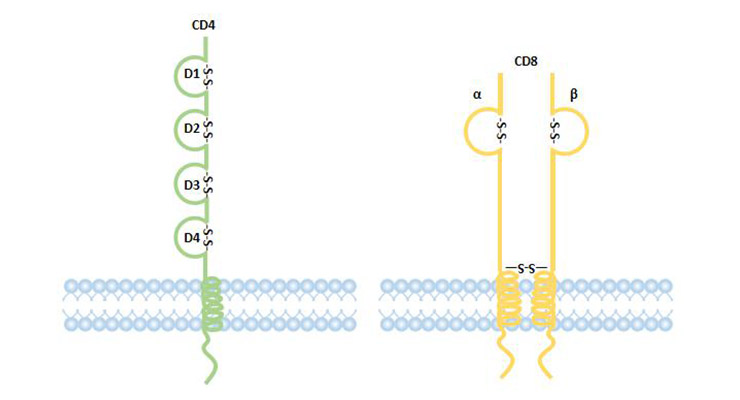
Figure 2 Structure of CD4 and CD8 coreptor
They are classic costimulatory molecules. Structurally, both are highly homologous and are homologous dimers formed by the joining of two polypeptide chains.
CD28: It is expressed in almost all CD4+ T cells, 50% CD8+ T cells. Plasma cells and partially activated B cells are also expressed.
CTLA-4: It expressed only on activated T cells.
The natural ligand of CD28 and CTLA-4 is B7 (CD80/CD86).
CD28-B7 (CD80/CD86) delivers a second signal of T cell activation (co-stimulatory signal).
CTLA-4 -B7 (CD80/CD86) gives activated T cell inhibition signal.
It belongs to the type Ⅱ transmembrane protein, mainly expressed in the activation of CD4 + T cells and CD8 + T cell surface. CD40L interacts with CD40 on the surface of B cells, providing synergistic stimulation signals to promote B cell proliferation, differentiation, antibody synthesis, and thymus-dependent antigen (TD-Ag)-induced immune responses.
Through these receptors, the virus can selectively infect a subset of T cells. For example, HIV can cause AIDS by infecting helper T cells (CD4+ cells).
Interleukin receptors, integrin receptors, transferrin receptors, and the like.
B cells are antigen-specific lymphocytes derived from hematopoietic tissue. Unlike T cells, B cells differentiate and mature in the bone marrow. After being stimulated by antigen, B lymphocytes are first transformed into plasmablasts, and then differentiated into plasma cells, produce and secrete immunoglobulins (antibodies), and participate in humoral immunity. Its function is to produce antibodies, present antigens, and secrete intracellular factors involved in immune regulation [16]. The detection of B cells and their subsets is an important indicator in the study of autoimmune diseases and immune regulation disorders in diseases.
It is an immunoglobulin embedded in the lipid molecules of cell membrane, and is expressed in mature B cells and most B cell tumor cells. Mature B cells mainly express mIgM and mIgD, and their contents also reflect the degree of cell maturation: IgM>IgD is more immature, IgD> IgM is more mature.
BCR is the most characteristic surface marker of B cells. It can bind to corresponding antigens specifically, and it can also bind to anti-immunoglobulin antibodies specifically.
The complement receptor is expressed on the surface of mature B cells, mainly the C3 receptor.
Complement receptor 1 (CR1), also known as C3b receptor or C3b/C4b receptor, binds to C3b and C4b and promotes B cell activation.
Complement receptor 2 (CD21), an important marker of B cells, also known as C3d receptor, is also a receptor for Epstein-Barr virus.
Natural killer cells are cells with large particles in the cytoplasm. The exact source of NK cells is not well understood and is generally thought to be derived directly from the bone marrow. Activated NK cells exert antiviral [17], anti-tumor [18] and immunoregulatory effects by secreting cytokines and cytotoxicity [19].
At present, CD3-CD16+CD56+ is often used as its typical symbol.
CD16 (FcγRIII), CD56 (NCAM-1), CD94, CD158 (KIR), and CD161 (NKR-P1A) are often selected for NK cell function analysis.
In summary, lymphocyte subsets can be distinguished by cell surface markers, as shown in table 2.
In summary, lymphocyte subsets can be distinguished by cell surface markers, as shown in table 2. Lymphocyte subsets can be distinguished by cell surface markers, as shown in table 2.
Table 2 Comparison of T, B and NK cell surface markers
| Surface marker | T cells | B cells | NK cells |
|---|---|---|---|
| Surface membrane immunoglobulin (SmIg) | - | + | - |
| TCR | + | - | - |
| CD2 | + | - | + |
| CD3 | + | - | - |
| CD19, CD20 | - | + | - |
| CD16, CD56 | - | - | + |
| Complement receptor (CR) | - | + | part |
| Fc receptor | - | + | + |
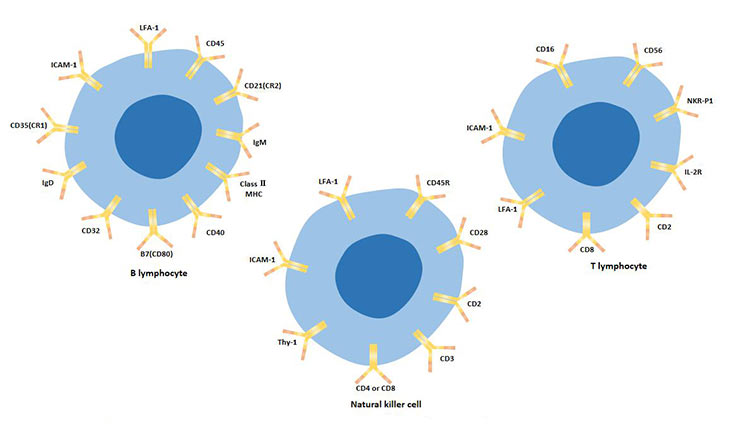
Figure 3 The cellular surface markers of T lymphocytes, B lymphocytes, and natural killer cells
It includes indirect immunofluorescence and direct immunofluorescence techniques.
The process of indirect immunofluorescence: First, the cells to be detected are treated with an unlabeled antibody to form a complex with a specific antigen, and then fluorescently labeled antibodies of anti-antibodies were used to detect the presence of specific antigens in the cells, which had fluorescence enhancement effect.
B cell surface markers-SmIg can be detected by indirect immunofluorescence.
This method is characterized by strong specificity and can accurately determine the distribution of fluorescence in tissues or cells by microscopic observation.
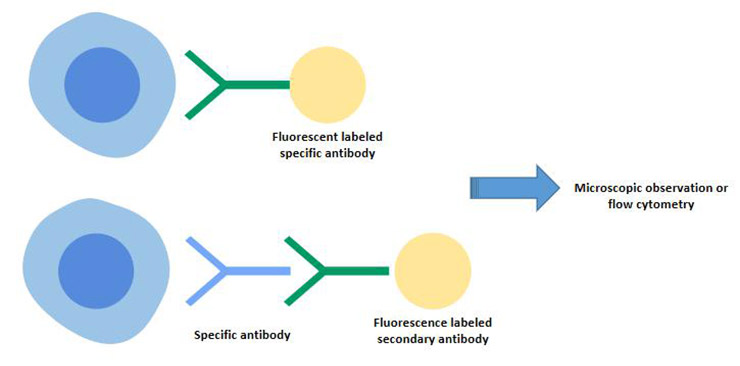
Figure 4 Indirect immunofluorescence and direct immunofluorescence techniques
This method typically employs an enzyme-linked immunoassay, such as the biotin-streptomycin system. Using a microscope, cells expressing brownish yellow color were judged to be CD antigen-positive cells.
Red blood cells labeled with the corresponding antibodies are used as indicators to mix with the cells to be detected.
Sheep red blood cells (SRBC) form spontaneous rosettes with human lymphocytes (E rosetta) [20] [21]. Therefore, it can be used to detect SRBC receptors on T cells. The principle of this method is to use the SRBC receptor on the surface of T cells to form a rosette with SRBC.
The method is simple and the reagent is relatively cheap. It is a classical method for T lymphocyte detection. But its disadvantage is that the result is not very stable, subject to personal subjective influence.
The analysis of lymphocyte subsets can reveal the balance of immune function in patients.
The immune function of the body is closely related to the occurrence and development of malignant tumors.
The T lymphocyte subsets were abnormal in the peripheral blood of patients with tumors. The CD3+ cells and CD4+ cells in the patients were significantly reduced, while the CD8+ cells were significantly increased. This immunosuppressive state reduces the patient's ability to recognize and kill mutant cells.
Lymphocyte subsets are often disturbed in patients with autoimmune diseases. Changes in the proportion of lymphocytes are closely related to the occurrence and development of autoimmune diseases [22].
An increase in the ratio of CD4+/CD8+ leads to autoimmune disease [23].
Lymphocyte surface markers can also be used to study the process and function of lymphocyte differentiation. Changes in the number and function of different groups of lymphocytes may also occur in the occurrence of various diseases. For example, CD4+T cells are significantly reduced or even deficient in AIDS patients.
Therefore, the detection of lymphocyte subsets is of great significance in controlling the occurrence and development of some diseases, understanding the pathogenesis and guiding clinical treatment.
References
[1] Niu J, Chang Y, Lu X, et al. Effect of dendritic cell vaccine therapy on lymphocyte subpopulation in refractory primary brain tumor [J]. Indian J Cancer, 2016, 52(4): 587 -589.
[2] Hsieh CT, Luo YH, Chien CS, et al. Induced pluripotent stem cellconditioned medium suppressed melanoma tumorigenicity through the enhancement of Natural-Killer cellular immunity [J]. J Immunother, 2016, 39(4): 153-159.
[3] Lisse I M, Qureshi K, Poulsen A, et al. T-lymphocyte subsets and eosinophil counts in acute and convalescence chickenpox infection: a household study in Guinea- Bissau [J]. Journal of Infection, 2005, 50(2): 125-129.
[4] Yushchuk N D, Gadzhikulieva M M, Balmasova I P, et al. The role of immune factors in the progression of chronic kidney diseases in HIV infection [J]. Ter Arkh, 2015, 88(3): 56-61.
[5] Bravo Soto JA, Esteban De La Rosa RJ, Luna Del Castillo JD, et al. Effect of mycophenolate mofetil regimen on peripheral blood lymphocyte subsets in kidney transplant recipients [J]. Transplant Proc, 2003, 35(4): 1355-1359.
[6] Cho JH, Yoon YD, Jang HM, et al. Immunologic monitoring of T-Lymphocyte subsets and Hla-Dr-Positive monocytes in kidney transplant recipients: a prospective, observational cohort study [J]. Medicine (Baltimore), 2015, 94(44): e1902.
[7] Papamichail M. T and B Lymphocytes: Origins, Properties and Roles in Immune Responses [J]. Immunology, 1975, 28(1).
[8] Yasutomo K. The cellular and molecular mechanism of CD4/CD8 lineage commitment [J]. Journal of Medical Investigation Jmi, 2002, 49(1-2): 1.
[9] Godfrey D I, Macdonald H R, Kronenberg M, et al. NKT cells: what's in a name? [J]. Nature Reviews Immunology, 2004, 4(3): 231-237.
[10] Yamaguchi Y, Ohshita A, Kawabuchi Y, et al. Adoptive immunotherapy of cancer using activated Autologous lymphocytes-current status and new strategies [J]. Human Cell, 2010, 16(4): 183-189.
[11] Brigl M, Brenner M B. CD1: Antigen Presentation and T Cell Function [J]. Annual Review of Immunology, 2004, 22(1): 817-890.
[12] Rogge L, Barberis - Manino L, Biffi M, et al. Selective expression of an IL - 12 receptor component by human T helper 1 cells [J]. J Exp Med, 1997 , 185: 825-831.
[13] Xu D,Chan WL,Leung BP, et al. Selective expression and functions of IL - 18 recptor on T helper (Th) type 1 but not Th2 cells [J ]. J Exp Med,1998 ,188 (8): 1485-1492.
[14] Mosmann TR, Cherwinski H, BondMW, et al. Two types of murine helper T cell clone. I. Defination according to profiles of lymphokine activities and secreated proteins [J]. J Immunol, 1986 , 136: 2348-2357.
[15] Appay V, Lier R A W V, Sallusto F, et al. Phenotype and function of human T lymphocyte subsets: Consensus and issues [J]. Cytometry, 2008, 73A(11): 975-983.
[16] Yu Q M, Yu C D, Ling Z Q. Elevated Circulating CD19(+) Lymphocytes Predict Survival Advantage in Patients with Gastric Cancer [J]. Asian Pacific journal of cancer prevention, 2012, 13(5): 2219-2224.
[17] Aura M, Marcel C G, Andrea V, et al. Priming of NK Cell Anti-Viral Effector Mechanisms by Direct Recognition of Human Cytomegalovirus [J]. Frontiers in Immunology, 2013, 0-4.
[18] Krzywinska E, Allende-Vega N, Cornillon A, et al. Identification of Anti-tumor Cells Carrying Natural Killer (NK) Cell Antigens in Patients With Hematological Cancers [J]. EBioMedicine, 2015, 2(10): 1364-1376.
[19] Galazka G, Jurewicz A, Domowicz M, et al. HINT1 peptide/Hsp70 complex induces NK-cell-dependent immunoregulation in a model of autoimmune demyelination [J]. Eur J Immunol, 2014, 44(10): 3026-3044.
[20] Lay W H, Mendes N F, Bianco C, et al. Binding of sheep red blood cells to a large population of human lymphocytes [J]. Nature, 1971, 230(5295): 531-2.
[21] Brain P, Gordon J. Rosette formation by peripheral lymphocytes. II. Inhibition of the phenomenon [J]. Clinical & Experimental Immunology, 1971, 8(3): 441-9.
[22] Gopalakrishnan S, Sen S, Adhikari J S, et al. The role of T-lymphocyte subsets and interleukin-5 blood levels among Indian subjects with autoimmune thyroid disease [J]. Hormones (Athens, Greece), 2010, 9(1): 76-81.
[23] Harrington L E, Hatton R D, Mangan P R, et al. Interleukin 17–producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages [J]. Nature Immunology, 2005, 6(11): 1123-1132.


Comments
Leave a Comment