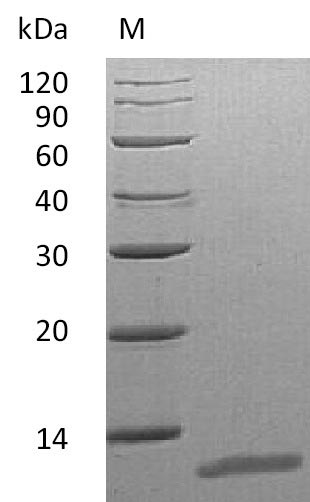
Purity
≥ 95% as determined by SDS-PAGE.
Endotoxin
Less than 1.0 EU/μg as determined by LAL method.
Activity
The ED50 as determined in a cell proliferation assay using TF?1 human erythroleukemic cells is less than 2 ng/ml.
Alternative Names
Il13; Il-13; Interleukin-13; IL-13; T-cell activation protein P600
Species
Mus musculus (Mouse)
Expression Region
22-131aa
Complete Sequence
PVPRSVSLPLTLKELIEELSNITQDQTPLCNGSMVWSVDLAAGGFCVALDSLTNISNCNAIYRTQRILHGLCNRKAPTTVSSLPDTKIEVAHFITKLLSYTKQLFRHGPF
Protein Length
Full Length of Mature Protein
Buffer
Lyophilized from a 0.2 μm filtered 20 mM PB, 150 mM NaCl, pH 7.2
Reconstitution
We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20°C/-80°C. Our default final concentration of glycerol is 50%. Customers could use it as reference.
Storage Condition
Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid
repeated freeze-thaw
cycles.
Shelf Life
The shelf life is related to many factors, storage state, buffer ingredients, storage
temperature
and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized
form is 12 months at -20°C/-80°C.
Lead Time
Basically, we can dispatch the products out in 5-10 working days after receiving your orders. Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.
Notes
Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
Datasheet & COA
Please contact us to get it.