Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
| Code | CSB-MP891791HU |
| Abbreviation | Recombinant Human TNFSF18 protein, partial (Active) |
| MSDS | |
| Size | $102 |
| Order now | |
| Image |
|
| Have Questions? | Leave a Message or Start an on-line Chat |

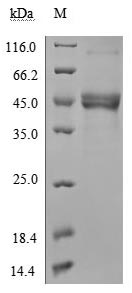
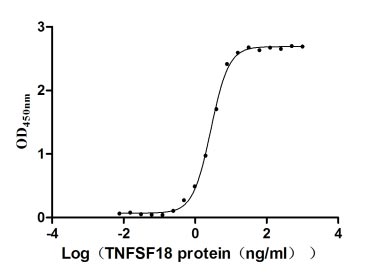
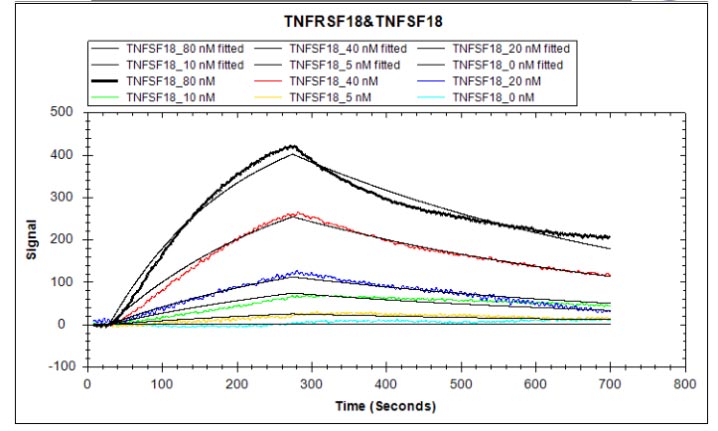
TNFSF18 (GITRL) plays a critical role in T-cell costimulation and regulatory T-cell homeostasis, making it a key target for immuno-oncology and autoimmunity research. This recombinant human TNFSF18 (aa 72–199), bearing an N-terminal hFc-Flag tag and expressed in mammalian cells, demonstrates robust receptor engagement with an EC50 of 2.565–2.940 ng/ml against immobilized TNFRSF18 in functional ELISA, and a KD of 38.5 nM by LSPR, confirming signaling-competent conformation suitable for NF-κB pathway activation studies and T-cell costimulation assays. The sub-nanogram EC50 supports use as a coating antigen or positive control in antibody development campaigns, while endotoxin levels below 1.0 EU/μg minimize LPS-driven artifacts in sensitive primary T-cell proliferation and activation experiments. Purity exceeding 90% by SDS-PAGE, combined with this stringent endotoxin profile, satisfies the quality criteria typical for cell-based functional assays and in vivo tumor biology models.
There are currently no reviews for this product.

