Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
| Code | CSB-MP013481MO |
| Abbreviation | Recombinant Mouse Mapt protein (Active) |
| MSDS | |
| Size | $138 |
| Order now | |
| Image |
|
| Have Questions? | Leave a Message or Start an on-line Chat |

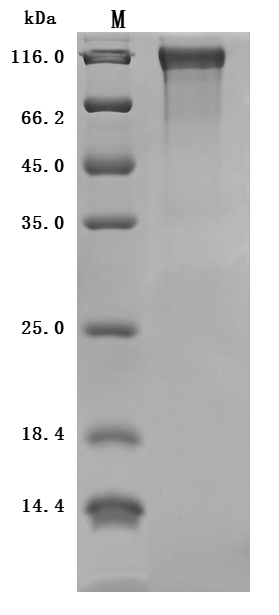
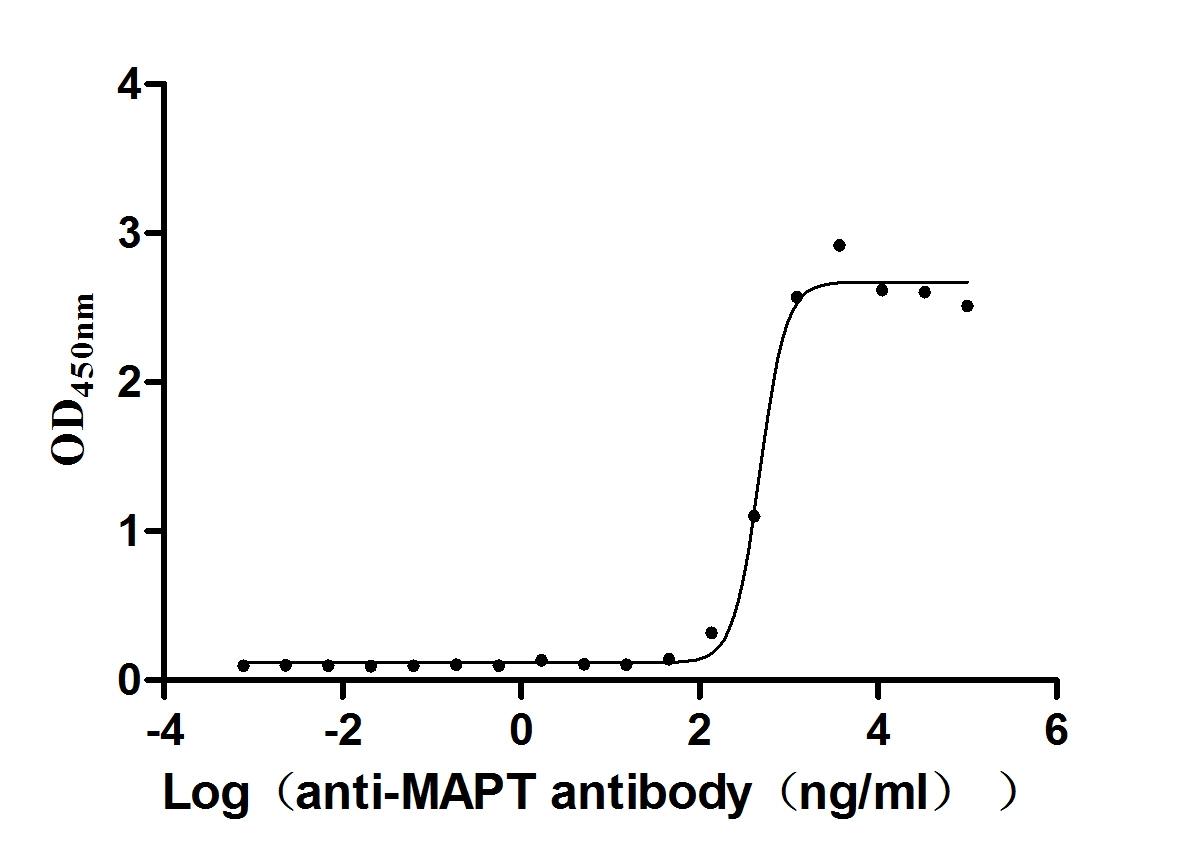
Tau protein plays a central role in microtubule stabilization and is the principal component of neurofibrillary tangles in neurodegenerative disease models, making functionally validated recombinant forms essential for mechanistic studies. This full-length mouse Mapt (residues 2–733) is expressed in mammalian cells with an N-terminal 10×His tag positioned outside the microtubule-binding repeat domains, preserving native folding and polymerization competence critical for cytoskeletal dynamics assays and structural studies such as cryo-EM. Functional ELISA confirms binding activity against an anti-MAPT recombinant antibody with an EC50 of 436.1–518.6 ng/mL, supporting direct use in protein-protein interaction assays and as a reliable positive control for Western blot or immunofluorescence-based antibody validation. Purity exceeding 90% by SDS-PAGE combined with endotoxin levels below 1.0 EU/μg satisfies the criteria typical for cell-based tau aggregation experiments and in vitro tubulin polymerization kinetics measurements.
There are currently no reviews for this product.

