Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
Recently, CDH17 has rapidly gained attention as a promising target for gastrointestinal cancers. In 2025, two significant BD transactions have drawn widespread interest: in October, Hansoh Pharma licensed the ex-Greater China rights of its CDH17 ADC (HS-20110) to Roche, with an upfront payment of $80 million and a potential total deal value of up to $1.53 billion. In January, Lepu Biopharma also granted the overseas rights of its CDH17 ADC (MRG007) to ArriVent, with a total transaction value exceeding $1.2 billion. Internationally, Multitude Therapeutics' CDH17 ADC (AMT-676) has entered Phase I clinical trials, and its trial design was presented at the 2025 ASCO conference. Domestically, Yilinked Therapeutics' YL217 has also progressed to the clinical stage. These advancements indicate that the CDH17 target is accelerating its transition from basic research into clinical applications.
Cadherins are a large superfamily of cell adhesion molecules. Cadherins form a superfamily with at least six subfamilies: classical cadherin (eg. E-cadherin and N-cadherin), atypical cadherin, desmocollins, desmogleins, protocadherin type, and others. Cadherins act as Ca2+-dependent transmembrane glycoproteins that mediate cell-cell adhesion, which are involved in a variety of signaling including Wnt/β-catenin, PI3K/Akt, Rho GTPase, NF-κB, and other pathways.
Many studies suggested that abnormal expression of cadherin is associated with various diseases, such as asthma [4], chronic periodontitis [5], atherosclerosis [6], and diabetes [7] etc. On top of that, cell adhesion molecules also play key roles in tumor suppression and cell migration. For example, E-cadherin is considered as an important tumor suppressor gene. N-cadherin functions mainly as a promoter of tumors [8-10]. Therefore, Cadherins may be served as therapeutic targets to treat allergies, autoimmune diseases, and tumors.
Cadherin-17 (corresponding to CDH17), also called liver–intestine (LI) cadherin or human peptide transporter-1 (HPT-1), is a newly discovered unique member of the cadherin superfamily [11]. CDH17 was originally cloned from rat liver in 1994. The human CDH17 gene is located on chromosome 8q22.1, which encodes LI-cadherin protein. CDH17 has similarity to the classic cadherins but is structurally distinct: i) CDH17 consists of seven cadherin repeats within the extracellular domain, and classic cadherins have five; ii) AAL-sequence motif is found, instead of the HAV-motif in the cell adhesion recognition site of the extracellular cadherin-1 (EC-1); iii) CDH17 has only 20 amino acids in the cytoplasmic domain, whereas classic cadherins have a highly conserved cytoplasmic domain consisting of 150-160 amino acids (Figure 1) [12-14].
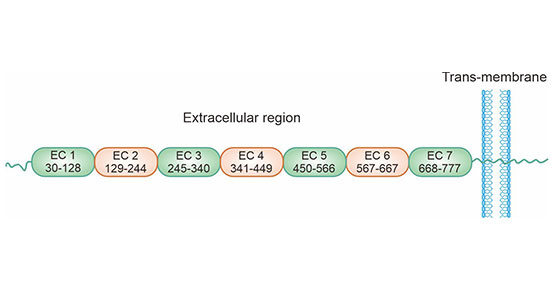
Figure 1. CDH17 structure [14]
CDH17 expression is highly restricted to epithelial cells in the small intestine and colon of both embryonic and adult mice and human. Notably, its expression is not detectable in the liver. CDH17 expression is known to be regulated by caudal-related homeobox transcription factor (CDX2) [12-14]. CDX2 has been reported as a tumor suppressive factor in colon cancer. It has been reported that CDH17 is a highly specific and more sensitive marker than CK20 and CDX2. In addition, CDH17 expression enhanced integrin activation and signaling, which is correlated with a moderately increased adhesion. Accumulated evidence has shown that CDH17 is overexpressed in many gastric cancer cells (GICs) [15-18].
CDH17 plays a crucial role in the cell adhesion process, similar to cadherin. β-catenin has essential roles in cell adhesion, but there is no direct association between CDH17 and β-catenin network [19-23]. CDH17 might perform its adhesion function by directly connecting to the cell scaffold. The precise biological function of CDH17 is still a matter of debate. To date, CDH17 has been identified in various tumors, including gastric, colorectal, hepatic, pancreatic and bile duct cancers. In a nutshell, CDH17 represents a previously unappreciated biomarker that might function as an independent adverse prognostic factor or therapeutic target for GIC patients [21-23].
Studies have revealed that the dysfunction of CDH17 is linked to the peripheral infiltration and metastasis of tumor cells, which has implications for tumor recurrence and patient survival. However, the mechanism of CDH17 in tumors has not been elucidated yet.
In gastric cancer, CDH17 overexpression dissociates the E-cadherin/catenin complex [23] that are involved in the cancer development via the Wnt signaling pathway [24-25]. In addition, CDH17 upregulation has influences on GSK3 activity, inhibiting the formation of β-catenin-AXIN-APC-GSK3 complex. Then increased β-catenin can bind to LEF/TCF, inducing Cyclin D1 expression, thus promoting cell proliferation and inhibiting apoptosis (Figure 2) [26]. It has been suggested that aberrant Galectin-3 in gastric cancer mediates CDH17 expression, but the exact regulatory mechanism needs to be further investigated [27-28].
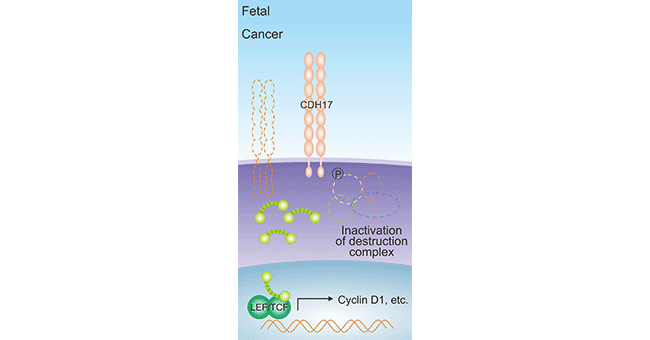
Figure 2. CDH17 induces Cyclin D1 expression [26]
In addition, up-regulated CDH17 expression enhances the activity of MMP-2 and MMP-9, facilitating tumor cell metastasis [17]. Plus, CDH17 could be enhanced by Ras/Raf/MEK/ERK signaling pathway that plays an important role in the proliferation of gastric cancer cells (Figure 3) [29]. In hepatocellular carcinoma, down-regulated CDH17 expression leads to inactivation of the Wnt/β-catenin signaling pathway and inhibition of tumor growth. Thus, it is hypothesized that targeting CDH17 might promote apoptosis of tumor cells via blocking Wnt signaling pathway [30].
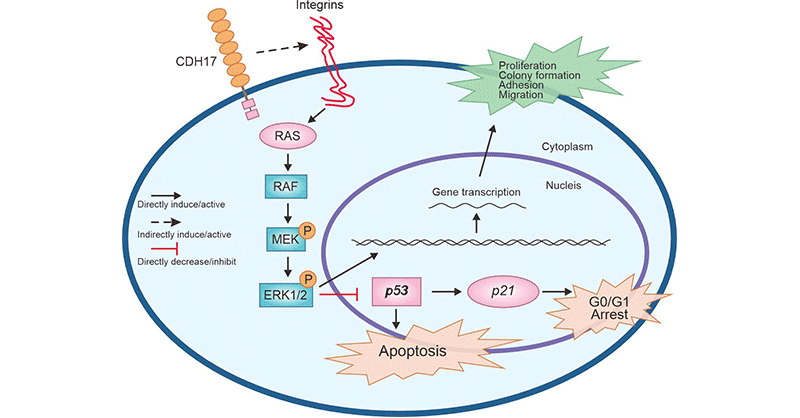
Figure 3. CDH17 mediates the Ras/Raf/MEK/ERK signaling pathway [29]
CDH17, as a novel cadherin, plays an important role in cell-cell adhesion and cell recognition. Dysregulation of intercellular adhesion and motility is one of the important mechanisms of tumor development. In recent years, there has been a growing focus on exploring the connection between CDH17 and tumors, particularly in gastrointestinal cancers like gastric, liver, colon, and pancreatic cancers.
CDH17 was first found to be overexpressed in gastric cancer. A clinical study from 71 gastric cancer patients revealed that CDH17 expression was positively correlated with the histological type, tumor invasion, and lymph node metastasis of gastric cancer. In the BGC-823 cell line, CDH17 overexpression promoted the proliferation, invasion, and migration of gastric adenocarcinoma. In MKN-45 gastric cancer cells, CDH17 downregulation curbed the proliferation, invasion, and adhesion of cells; NF-κB signaling pathway was inhibited and its downstream protein (VEGF-C and MMP-9) were reduced [31-33].
CDH17 is also related to liver cancer. The analysis of 57 cases by RT-PCR showed that CDH17 in liver cancer tissues was significantly higher than that in normal groups, indicating that CDH17 may be used as a tumor marker for early diagnosis of liver cancer [34]. A study of 34 cases of intrahepatic cholangiocarcinoma found that CDH17 was associated with the degree of tumor differentiation and vascular invasion. CDH17 knockdown promoted MTF-1 and PLGF expression, which in turn induced tumor angiogenesis [11, 35]. Another study found that splice isoforms and gene polymorphisms of CDH17 increase the risk of hepatocellular carcinogenesis [26].
A study of 45 colon cancer specimens found that down-regulation of CDH17 was significantly associated with colon cancer progression and lymph node metastasis. The consistent expression has been observed between CDH17 expression in colon cancer tissues at the primary site and metastatic sites. Therefore, CDH17 shows potential as a marker for detecting the metastasis of colon cancer [22, 36].
One study demonstrated a relationship between CDH17 immunoreactivity in esophageal adenocarcinoma and clinical characteristics of patients. The data found that there was a higher CDH17 immunoreactivity in well-differentiated esophageal cancer tissues, while the CDH17 was low or insignificant in low-differentiated esophageal adenocarcinoma tissues [37]. In esophageal squamous cell carcinoma ESCC, CDH17 CpG island methylation was enhanced and CDH17 was decreased [38-39]. Therefore, CDH17 could be a new target for esophageal cancer treatment.
In pancreatic cancer, CDH17 expression was higher in highly differentiated pancreatic ductal carcinoma than in those with poorly differentiated carcinomas. Kaplan-Meier analysis indicated that high CDH17 expression was associated with prognostic survival [40]. Low CDH17 expression was associated with tumor dedifferentiation. Additional research is required to fully understand the correlation between CDH17 and clinicopathological features of pancreatic cancer, as there are few of relevant studies.
CDH17 is also found in high levels in other types of cancer, like ovarian cancer and intraductal papillary mucinous neoplasm (IPMN). CDH17 is highly expressed in ovarian epithelial carcinoma, which was associated with poor prognosis [41]. CDH17 also plays a role in causing intestinal-type differentiation in IPMN [42]. Overall, CDH17 may be helpful in diagnosing and predicting the progression and recurrence of certain cancers such as gastric, hepatocellular, and colorectal cancers.
Several CDH17-based clinical agents are currently under development (Table 1), mainly for the treatment of gastrointestinal cancers. Among them, three are three bispecific antibodies: ARB-202 (CDH17 x CD3), ARB-001.T (CDH17 x CD3), BI-905711 (CDH17 x DR5/TRAIL-R2); one CAR-T drug CHM-2101; and two monoclonal antibodies. Surgery accompanied by chemotherapy and radiotherapy remains the first option for gastrointestinal tumors, but they aren't always very effective.
Tumor-targeted immunotherapy presents a promising new approach to treating cancer that is highly effective, safe, and low-toxic. This innovative strategy has the potential to revolutionize the current treatment landscape. Notably, recent research findings have demonstrated that inhibiting CDH17 can effectively impede tumor growth. As such, CDH17, as a newly discovered liver-intestine cadherin, is becoming a potential target for gastrointestinal tumors!
| Drugs | Target | Mechanism of action | Drug Type | Indications | Institutes | R&D status |
|---|---|---|---|---|---|---|
| ARB-202 | CDH17; CD3 | Immunomodulators. CDH17 modulator; CD3 modulator |
Bispecific antibodies | Bile duct cancer. Colorectal cancer. Gastrointestinal tumors; gastric cancer; biliary tract tumors; pancreatic cancer; liver cancer; pancreatic alveolar cancer |
Arbele | Clinical Phase 1 |
| BI-905711 | DR5; CDH17 | DR5 agonist. CDH17 modulators |
Bispecific antibodies | Diffuse large B-cell lymphoma. Gastric cancer. Bile duct cancer; gastrointestinal tract tumor; pancreatic cancer; esophageal cancer; bile duct cancer |
C.H. Boehringer Sohn AG & Co. KG; Boehringer Ingelheim (China) Investment Co. Boehringer Ingelheim (China) Investment Co. Boehringer Ingelheim GmbH |
Clinical Phase 1 |
| Anti-CDH17 mAbs (ProAlt) | CDH17 | CDH17 modulator | Monoclonal antibodies | Metastatic colorectal cancer; metastatic melanoma | Proclara Biosciences, Inc. | Preclinical |
| ARB-001. | CDH17 | CDH17 antagonist | Bispecific antibodies | Gastrointestinal tumors | Arbele | Preclinical |
| ARB-001. | CD3; CDH17 | CDH17 modulator | Bispecific antibodies | Colorectal cancer; liver cancer. Stomach cancer |
Arbele | Preclinical |
| ARB-201 | CDH17 | CDH17 antagonist; CD3 inhibitor | Bispecific antibodies | Colorectal Cancer | Arbele | Preclinical |
| CHM-2101 | CDH17 | CDH17 regulator; gene transfer | CAR-T | Colorectal cancer. Gastrointestinal tumors. Neuroendocrine tumors |
/ | Preclinical |
| CHM-2301 | CDH17 | CDH17 modulator | CAR-NK | Solid tumors | / | Preclinical |
| Anti-CDH17-based antibody drug conjugates (ProAlt) | CDH17 | CDH17 modulator | ADC; monoclonal antibody | Colorectal Cancer | Protein Alternatives SL | Drug Discovery |
Table 1: Clinical drugs in development for CDH17
To fully support researchers and pharmaceutical companies in their research on CDH17 in gastrointestinal cancers, CUSABIO presents CDH17 active protein (Code: CSB-MP613267HU) to support your research on the mechanism of CDH17 or its potential clinical value. (click for the full list of CDH17 products: CDH17 Protein; CDH17 antibody).
● CDH17 Recombinant Protein
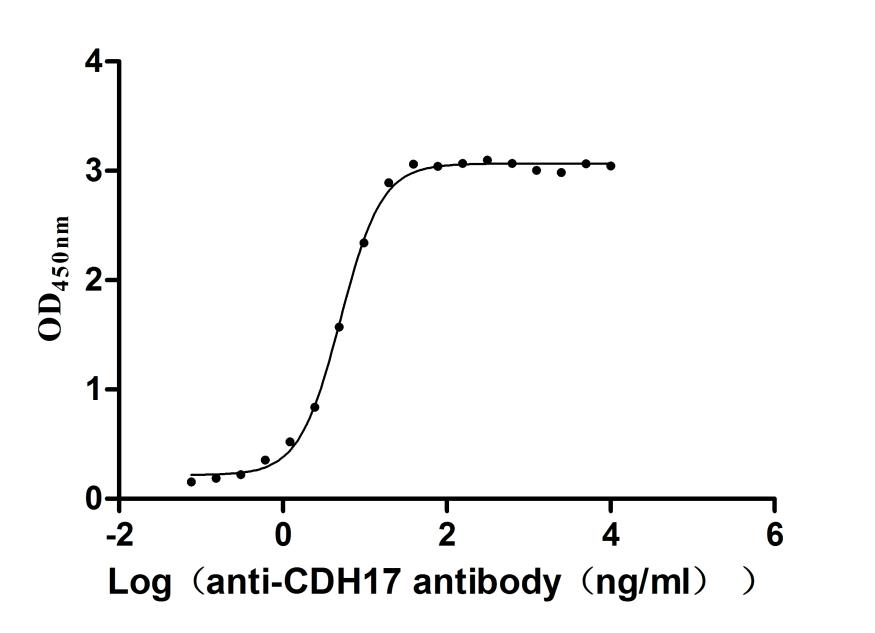
Recombinant Human Cadherin-17 (CDH17), partial (Active); CSB-MP613267HU
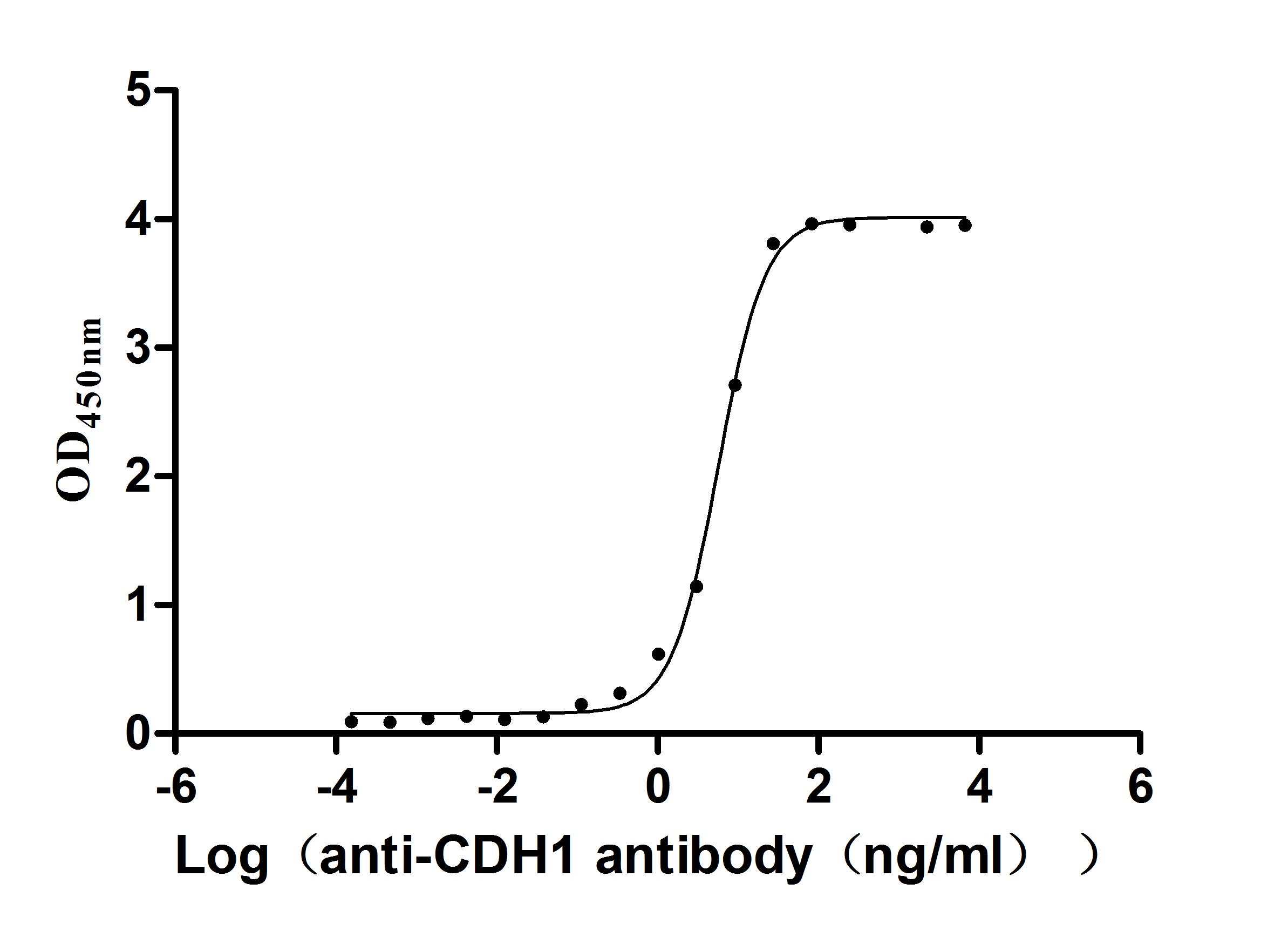
Recombinant Human Cadherin-1(CDH1),partial (Active); CSB-MP005034HU1
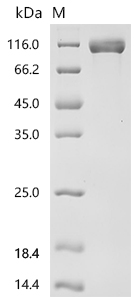
Recombinant Macaca fascicularis Cadherin 17 (CDH17), partial (Active); CSB-MP4664MOV

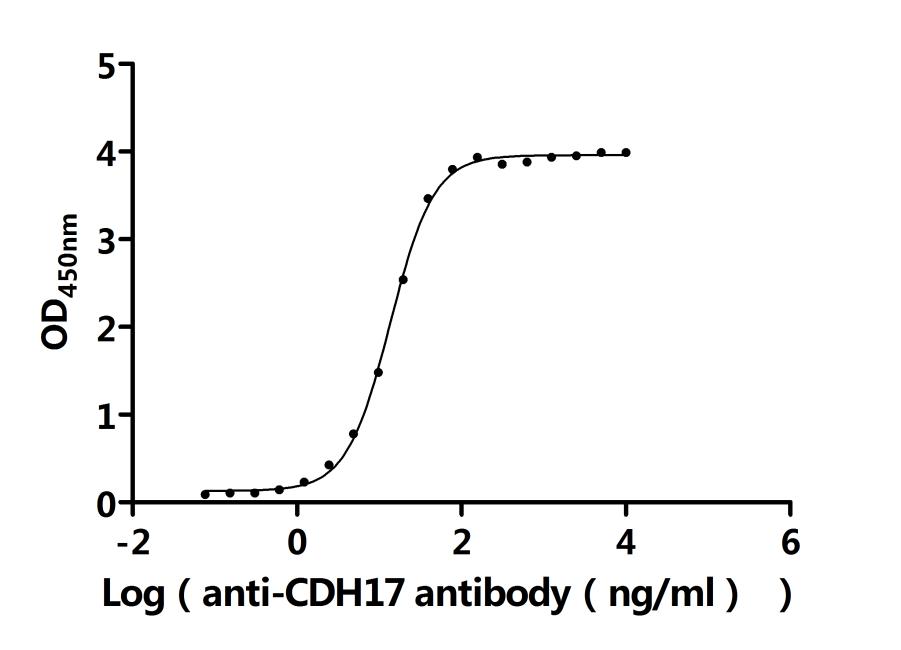
Recombinant Rat Cadherin-17 (Cdh17), partial (Active); CSB-MP005042RA
● CDH17 Antibody
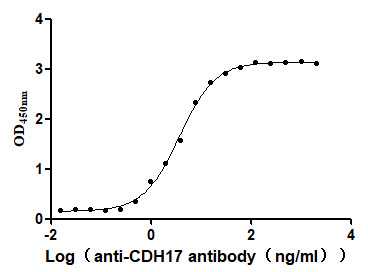
CDH17 Recombinant Monoclonal Antibody; CSB-RA613267MA3HU
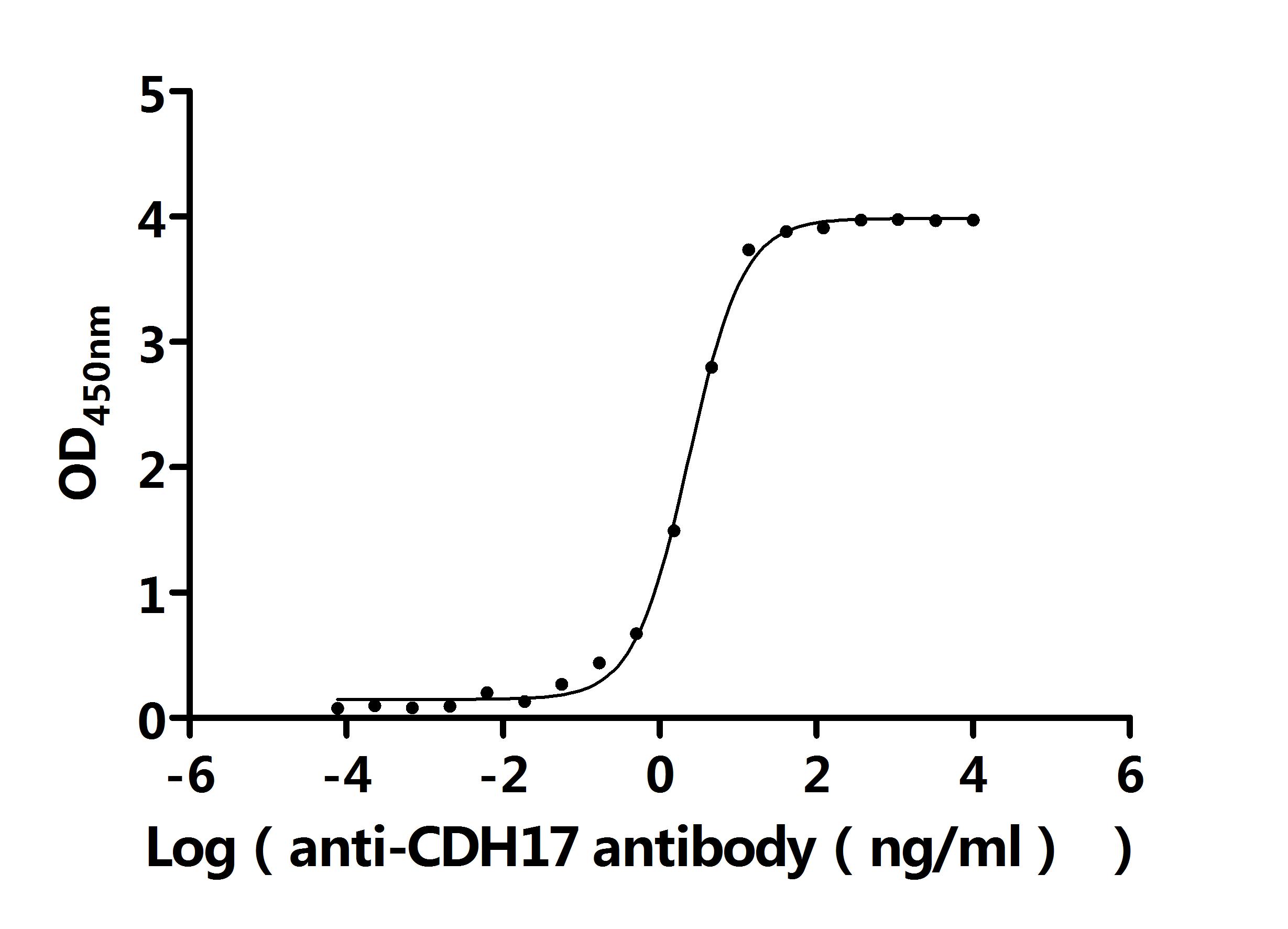
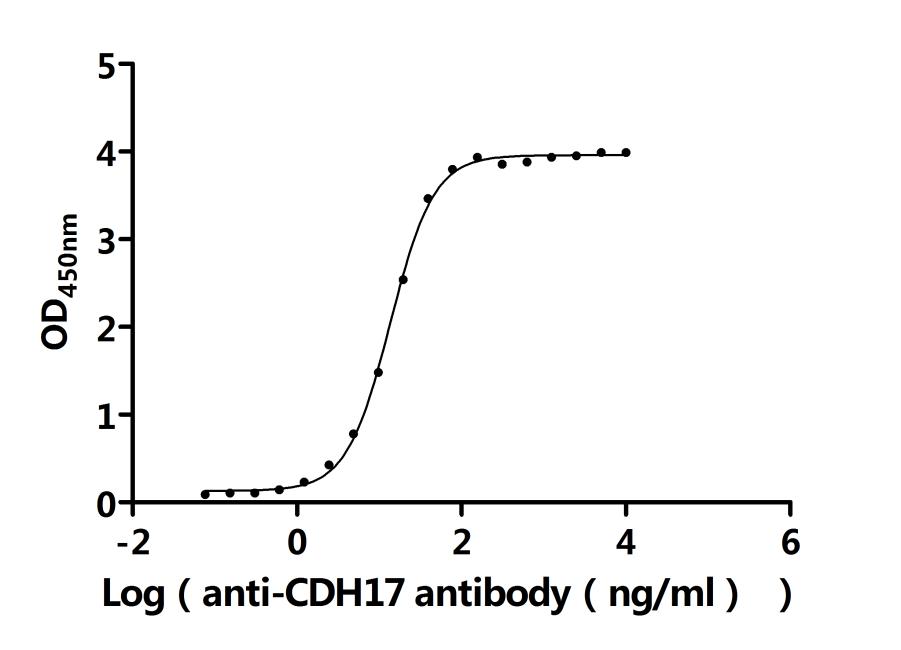
CDH17 Recombinant Monoclonal Antibody; CSB-RA613267MA1HU
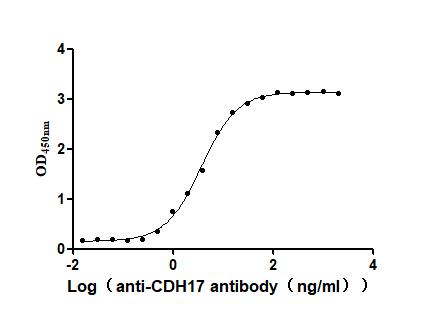
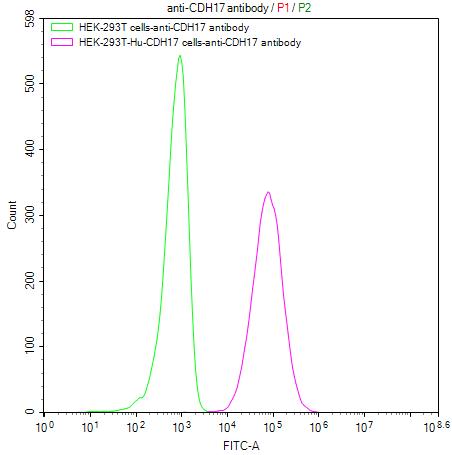
● CDH17 Stable Cell Line
References
[1] Hulpiau, Paco, Ismail Sahin Gul, and Frans Van Roy. "Evolution of cadherins and associated catenins. "The Cadherin Superfamily: Key Regulators of Animal Development and Physiology (2016): 13-37.
[2] Ma, Yi-Shih, et al. "Bisdemethoxycurcumin suppresses human osteosarcoma U-2 OS cell migration and invasion via affecting the PI3K/Akt/ NF-κB, PI3K/Akt/GSK3β and MAPK signaling pathways in vitro." Oncology Reports 48.6 (2022): 1-10.
[3] Yu, Weina, et al. "Cadherin signaling in cancer: its functions and role as a therapeutic target." Frontiers in oncology 9 (2019): 989.
[4] Türkeli, Ahmet, et al. "Anti-VEGF treatment suppresses remodeling factors and restores epithelial barrier function through the E -cadherin/β-catenin signaling axis in experimental asthma models." Experimental and Therapeutic Medicine 22.1 (2021): 1 -9.
[5] Ramesh, P. K. Immunolocalization of Gingival E-Cadherin Expression in Smokers and Non-Smokers with Chronic Periodontitis. diss. KSR Institute of Dental Science and Research, Tiruchengode, 2020.
[6] Harki, Olfa, et al. "Inhibition of Vascular Endothelial Cadherin Cleavage Prevents Elastic Fiber Alterations and Atherosclerosis Induced by Intermittent Hypoxia in the Mouse Aorta." International Journal of Molecular Sciences 23.13 (2022): 7012.
[7] Koziolek, Michael, et al. "Urine E-cadherin: a marker for early detection of kidney injury in diabetic patients." journal of clinical medicine 9.3 (2020 ): 639.
[8] Loh, Chin-Yap, et al. "The E-cadherin and N-cadherin switch in epithelial-to-mesenchymal transition: signaling, therapeutic implications, and challenges." Cells 8.10 (2019): 1118.
[9] Yu, Chong, et al. "The lncRNA ZNF667-AS1 Inhibits Propagation, Invasion, and Angiogenesis of Gastric Cancer by Silencing the Expression of N-Cadherin and VEGFA." Journal of Oncology 2022 (2022).
[10] Zhang, Jinyao, et al. "Clinical significance of ALDH1A1 expression and its association with E-cadherin and N-cadherin in resected large cell neuroendocrine carcinoma." Translational Oncology 19 (2022): 101379.
[11] Takamura, Masaaki, et al. "Involvement of liver-intestine cadherin in cancer progression." medical molecular morphology 46 (2013): 1-7.
[12] Ordóñez, Nelson G. "Cadherin 17 is a novel diagnostic marker for adenocarcinomas of the digestive system." Advances in anatomic pathology 21.2 (2014): 131-137.
[13] Gray, Michelle E., and Marcos Sotomayor. "Crystal structure of the nonclassical cadherin-17 N-terminus and implications for its adhesive binding mechanism." Acta Crystallographica Section F: Structural Biology Communications 77.3 (2021): 85-94.
[14] Caporuscio, Christian, et al. "Immunoaffinity enrichment LC-MS/MS quantitation of CDH17 in tissues." Bioanalysis 12.20 (2020): 1439- 1447.
[15] Horsfield, Julia, et al. "Cadherin-17 is required to maintain pronephric duct integrity during zebrafish development." Mechanisms of development 115.1-2 (2002): 15-26.
[16] Huang, Li-Ping, et al. "Up-regulation of cadherin 17 and down-regulation of homeodomain protein CDX2 correlate with tumor progression and unfavorable prognosis in epithelial ovarian cancer." International Journal of Gynecologic Cancer 22.7 (2012).
[17] Jiang, Xiao-jie, et al. "CDH17 alters MMP-2 expression via canonical NF-κB signalling in human gastric cancer." Gene 682 (2019): 92-100.
[18] Liu, Xinjian, et al. "Disruption of oncogenic liver-intestine cadherin (CDH17) drives apoptotic pancreatic cancer death." Cancer Letters 454 (2019): 204-214.
[19] Xia, Peng, et al. "Surface-Engineered Extracellular Vesicles with CDH17 Nanobodies to Efficiently Deliver Imaging Probes and Chemo -Photothermal Drugs for Gastric Cancer Theragnostic." Advanced Functional Materials (2022): 2209393.
[20] Xia, Peng, et al. "Surface-Engineered Extracellular Vesicles with CDH17 Nanobodies to Efficiently Deliver Imaging Probes and Chemo -Photothermal Drugs for Gastric Cancer Theragnostic." Advanced Functional Materials (2022): 2209393.
[21] Feng, Zijie, et al. "Potent suppression of neuroendocrine tumors and gastrointestinal cancers by CDH17CAR T cells without toxicity to normal tissues." Nature Cancer 3.5 (2022): 581-594.
[22] Pei, Xiao Meng, et al. "The diagnostic significance of CDH17-positive circulating tumor cells in patients with colorectal cancer." Expert Review of Molecular Diagnostics just-accepted (2023).
[23] Wu, Cunen, et al. "Interaction between Wnt/β-catenin pathway and microRNAs regulates epithelial-mesenchymal transition in gastric cancer." International journal of oncology 48.6 (2016): 2236-
[22] 6.
[24] Kaszak, Ilona, et al. "Role of cadherins in cancer-a review." international journal of molecular sciences 21.20 (2020): 7624.
[25] Wang, Qianwen, et al. "MICAL2 contributes to gastric cancer cell migration via Cdc42-dependent activation of E-cadherin/β-catenin signaling pathway ." Cell Communication and Signaling 20.1 (2022): 136.
[26] Lee, Nikki P., et al. "Role of cadherin-17 in oncogenesis and potential therapeutic implications in hepatocellular carcinoma. "Biochimica et Biophysica Acta (BBA)-Reviews on Cancer 1806.2 (2010): 138-145.
[27] Huang, Hsiang-Wei, et al. "Association between inflammation and function of cell adhesion molecules influence on gastrointestinal cancer development." Cells 10.1 (2021): 67.
[28] Maher, John, and David M. Davies. "CAR-Based Immunotherapy of Solid Tumours-A Survey of the Emerging Targets. "Cancers 15.4 (2023): 1171.
[29] Lin, Zhaohu, et al. "Targeting cadherin-17 inactivates Ras/Raf/MEK/ERK signaling and inhibits cell proliferation in gastric cancer." PLoS One 9.1 ( 2014): e85296.
[30] Wang, Yonggang, et al. "Anti-cadherin-17 antibody modulates beta-catenin signaling and tumorigenicity of hepatocellular carcinoma." ploS one 8.9 ( 2013): e72386.
[31] Motoshita, Junichi, et al. "Molecular characteristics of differentiated-type gastric carcinoma with distinct mucin phenotype: LI -cadherin is associated with intestinal phenotype." Pathology international 56.4 (2006): 200-205.
[32] Wang, Jin, et al. "Cadherin-17 induces tumorigenesis and lymphatic metastasis in gastric cancer through activation of NFκB signaling pathway." Cancer biology & therapy 14.3 (2013): 262-270.
[33] Qiu, Hai-bo, et al. "Targeting CDH17 suppresses tumor progression in gastric cancer by downregulating Wnt/β-catenin signaling." ploS one 8.3 (2013): e56959.
[34] Su, Min-Cheng, et al. "Cadherin-17 is a useful diagnostic marker for adenocarcinomas of the digestive system." Modern Pathology 21.11 (2008): 1379- 1386.
[35] Takamura, Masaaki, et al. "Loss of liver-intestine cadherin in human intrahepatic cholangiocarcinoma promotes angiogenesis by up-regulating metal- responsive transcription factor-1 and placental growth factor." International journal of oncology 36.1 (2010): 245-254.
[36] Harding, J. J., et al. "371P A phase Ia/b, open-label, multicentre study of the TRAILR2 agonist BI 905711 in patients (pts) with advanced gastrointestinal (GI) cancers." Annals of Oncology 33 (2022): S706.
[37] Panarelli, Nicole C., et al. "Tissue-specific cadherin CDH17 is a useful marker of gastrointestinal adenocarcinomas with higher sensitivity than CDX2 ." American journal of clinical pathology 138.2 (2012): 211-222.
[38] Shenoy, U. Sangeetha, et al. "Molecular implications of HOX genes targeting multiple signaling pathways in cancer." Cell biology and toxicology (2022) : 1-30. : 1-30.
[39] Ignatova, Ekaterina Olegovna, et al. "Clinical significance of molecular subtypes of gastrointestinal tract adenocarcinoma." World Journal of Gastrointestinal Oncology 14.3 (2022): 628.
[40] Liu, Xinjian, et al. "Disruption of oncogenic liver-intestine cadherin (CDH17) drives apoptotic pancreatic cancer death." Cancer Letters 454 (2019): 204-214.
[41] Huang, Li-Ping, et al. "Up-regulation of cadherin 17 and down-regulation of homeodomain protein CDX2 correlate with tumor progression and unfavorable prognosis in epithelial ovarian cancer." International Journal of Gynecologic Cancer 22.7 (2012).
[42] Karimi, S. S., T. Valyi-Nagy, and M. F. Gonzalez. "TTF-1 Immunoexpression in Primary Rectal Adenocarcinoma with Brain Metastasis. "American Journal of Clinical Pathology 156.Supplement_1 (2021): S63-S63.


Comments
Leave a Comment