Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
Microglia, the resident immune cells of the central nervous system (CNS), have emerged as key regulators of brain development, homeostasis, and disease pathogenesis. Once considered passive supportive cells, decades of research have revealed their extraordinary dynamicity and multifaceted roles in immune surveillance, inflammation, and neural circuit remodeling. The identification and validation of specific microglial markers have been instrumental in advancing our understanding of these cells, enabling precise characterization of their activation states and functional contributions in health and disease.
This article provides a comprehensive overview of microglial biology, including their origin, phenotypic plasticity, core functions, and the most reliable markers for their identification and characterization. We also highlight markers that distinguish resident microglia from peripheral macrophages and define distinct activation phenotypes.
Table of Contents
Microglia are the primary myeloid cells of the CNS, comprising approximately 10–20% of all glial cells in the adult brain and spinal cord [1,2]. A defining feature that distinguishes them from peripheral macrophages is their developmental origin: microglia arise from primitive yolk sac myeloid progenitors during early embryogenesis, migrate to the developing CNS, and self-renew locally throughout life, whereas peripheral macrophages derive from bone marrow hematopoietic stem cells [3].
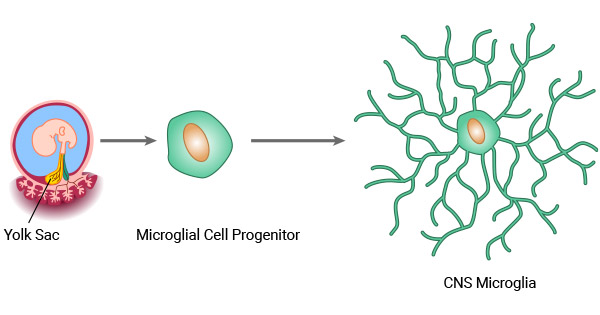
Figure 1. The origin of microglia [9]
In the healthy, uninjured brain, microglia exist in a homeostatic (M0) state characterized by a highly ramified morphology. Their small cell bodies extend fine, motile processes that continuously survey the extracellular microenvironment, detecting perturbations such as tissue damage, pathogen invasion, or neuronal dysfunction.
Upon activation, microglia undergo dramatic morphological and functional transformation. They retract their ramified processes, adopt an amoeboid or hypertrophic shape, and acquire enhanced phagocytic and migratory capabilities. Activated microglia migrate to sites of injury or infection, where they engulf and clear cellular debris, dead neurons, protein aggregates, and invading pathogens [4–8].
Microglia exhibit remarkable phenotypic plasticity, dynamically adjusting their gene expression and function in response to diverse microenvironmental cues. Historically, activated microglia have been categorized into two opposing phenotypes: the pro-inflammatory "classically activated" M1 phenotype and the anti-inflammatory "alternatively activated" M2 phenotype.
M1 polarization is typically induced by pathogen-associated molecular patterns (PAMPs) or pro-inflammatory cytokines such as interferon-γ (IFN-γ) and tumor necrosis factor-α (TNF-α). M1 microglia produce high levels of pro-inflammatory mediators including CD16, CD32, CD86, interleukin-1β (IL-1β), TNF-α, IL-6, and inducible nitric oxide synthase (iNOS). This phenotype plays a critical role in host defense against pathogens and the initiation of immune responses. However, excessive or prolonged M1 activation drives chronic neuroinflammation, which contributes to neuronal damage in numerous neurological disorders.
M2 microglia are alternatively activated and associated with inflammation resolution, tissue repair, and homeostasis maintenance. They are characterized by upregulated expression of markers such as CD163, CD206 (mannose receptor), and arginase-1 (Arg1). The M2 phenotype is further divided into three functionally distinct subsets:

Figure 2. Activated microglia markers of the M1 and M2 polarization spectrum—cellular and released [9]
Importantly, the traditional M1/M2 binary classification is now widely recognized as an oversimplification of microglial plasticity. In vivo, microglia exist along a continuous spectrum of activation states with overlapping gene expression profiles and mixed functional properties, shaped by tissue context, disease stage, and local microenvironmental signals [21]. This heterogeneity underscores the need for multiple complementary markers to define microglial phenotypes accurately.
Microglia are essential for CNS development, physiological homeostasis, and response to injury or disease [10]. In the healthy brain, they act as immune sentinels, continuously monitoring the microenvironment for signs of disturbance.
A core function of microglia is phagocytosis. They clear apoptotic neurons, cellular debris, and misfolded protein aggregates to maintain tissue integrity. During postnatal development, microglia play a critical role in neural circuit refinement by pruning excess synapses via complement-dependent pathways (e.g., C1q and C3), a process essential for normal cognitive function and synaptic plasticity [22].
Microglia also support neuronal survival and function by secreting neurotrophic factors such as brain-derived neurotrophic factor (BDNF) and insulin-like growth factor 1 (IGF-1). They contribute to maintaining the integrity of the blood-brain barrier and communicate bidirectionally with neurons, astrocytes, and oligodendrocytes to regulate overall brain homeostasis.
Dysregulated microglial activation is a hallmark of many neurological disorders, including Alzheimer's disease [11–13], multiple sclerosis [14], Parkinson's disease, amyotrophic lateral sclerosis, and ischemic stroke [15,16]. Depending on the context, microglia can exert both neuroprotective and neurotoxic effects, making them attractive therapeutic targets.
Microglial markers are indispensable tools for identifying, quantifying, and characterizing microglia in tissue samples and in vitro models. They enable researchers to distinguish microglia from other CNS cell types, assess their activation states, and study their functional roles in health and disease.
A key challenge in microglia research is distinguishing resident microglia from infiltrating peripheral macrophages, which share many common myeloid markers. Recent advances have identified highly specific markers that allow unambiguous identification of resident microglia in both mouse and human tissues.
CUSABIO provides a comprehensive range of high-quality antibodies and ELISA kits validated for detecting microglial markers across applications such as flow cytometry (FC), immunohistochemistry (IHC), and immunofluorescence (IF).
| Classification | Marker | Expression Profile | CUSABIO Antibodies | CUSABIO ELISA Kits |
|---|---|---|---|---|
| General Myeloid Markers (label microglia and other myeloid cells) | Iba1 | Selectively expressed in microglia and macrophages. Gold standard for IHC/IF visualization of microglia [17]. | Iba1 Antibody | Human Allograft inflammatory factor 1(AIF1) ELISA kit |
| CD11b | Expressed on all myeloid cells. Upregulated during microglial activation. Commonly used with CD45 to distinguish microglia from peripheral myeloid cells. | CD11b Antibody | Human Integrin αM, ITG/CD11b ELISA Kit | |
| CD68 | Lysosomal marker for macrophage lineage cells. Primarily labels microglia and perivascular macrophages in the CNS [18]. | CD68 Antibody | Human CD68 ELISA kit | |
| CD45 | Leukocyte common antigen. Microglia express intermediate levels (CD45int), while peripheral macrophages express high levels (CD45hi). | CD45 Monoclonal Antibody | / | |
| F4/80 | Mouse macrophage marker. Expressed on both microglia and peripheral macrophages. | / | / | |
| Highly Specific Microglial Markers (not expressed by peripheral macrophages) | P2RY12 | Exclusively expressed by resident microglia in the CNS. Downregulated upon activation. Involved in process surveillance and chemotaxis [23]. | / | / |
| TMEM119 | Transmembrane protein specific to microglia. Does not stain infiltrating monocytes or CNS-associated macrophages [24]. | / | Human TMEM119 ELISA Kit | |
| TREM2 | Expressed on microglia and some peripheral myeloid cells. Critical for phagocytosis and lipid metabolism. Upregulated in disease-associated microglia (DAM) [25]. | TREM2 Recombinant Monoclonal Antibody | Human TREM2 ELISA Kit | |
| SALL1 | Nuclear transcription factor specific to microglia. Distinguishes microglia from all other myeloid cells in humans and mice [26]. | / | / | |
| CX3CR1 | Expressed on microglia and some peripheral monocytes. Widely used in CX3CR1-GFP reporter mice. | CX3CR1 Antibody | Human CX3CR1 ELISA Kit | |
| Siglec-H | Mouse-specific microglial marker. Absent from CNS-associated macrophages and infiltrating monocytes. | / | / | |
| FCRLS | Highly specific to murine microglia. No human ortholog. | / | / | |
| M1 Activation Markers | CD16 | Involved in ADCC and pro-inflammatory responses. Upregulated in M1 microglia. | Fcgr3 Antibody | / |
| CD32 | Upregulated during M1 polarization and commonly used in FC to identify and quantify M1 microglia. | FCGR2B Antibody | Human FCGR2B ELISA kit | |
| CD86 | Upregulated on M1-phenotype microglia and associated with pro-inflammatory cytokine production. Frequently used as a marker for M1 polarization in experimental studies. | CD86 Antibody | Human soluble CD86 ELISA Kit | |
| iNOS | Upregulated in M1-phenotype microglia. A key marker for M1 microglial activation and neuroinflammation. | NOS2 Antibody | Human iNOS ELISA KIT | |
| IL-1β | Involved in the amplification of neuroinflammation. A hallmark of M1 polarization. Commonly used as a functional marker to assess M1 microglial activation. | IL1B Antibody | Human IL-1β ELISA Kit | |
| M2 Activation Markers | CD206 | Upregulated in M2-phenotype microglia and associated with anti-inflammatory and tissue repair functions. Widely used as a marker for M2 polarization in experimental studies. | MRC1 Antibody | Human MRC1 ELISA Kit |
| Arg1 | Highly expressed in M2-phenotype microglia and associated with the production of anti-inflammatory factors [19]. A key marker for M2 polarization and tissue healing processes. | Arg1 Antibody | Human Arg1 ELISA kit | |
| CD163 | Scavenger receptor for hemoglobin-haptoglobin complexes. Associated with anti-inflammatory functions [20]. | CD163 Recombinant Monoclonal Antibody | Human soluble CD163 ELISA Kit | |
| Ym1 | Highly expressed in M2-phenotype microglia and commonly used as markers for M2 polarization. Involved in tissue repair, immunomodulation, and anti-inflammatory responses. | / | / | |
| Ym2 | Involved in tissue remodeling, immunomodulation, and anti-inflammatory responses. | / | / | |
| Context-Dependent Markers | Ferritin | Iron storage protein. Levels increase with microglial activation and inflammation. | / | Human Ferritin ELISA Kit |
| Vimentin | Intermediate filament protein. Expressed during development and in activated microglia in injury and neurodegeneration. | VIM Antibody | Human vimentin ELISA Kit |
It is critical to recognize that no single marker is universally specific for all microglial states. Marker expression can vary significantly by species, brain region, developmental stage, and disease context. Therefore, a panel of complementary markers and functional assays should be used to comprehensively characterize microglial phenotypes and functions.
Q: Is Iba1 specific to microglia?
A: No. Iba1 is expressed by all myeloid cells, including microglia, macrophages, monocytes, and dendritic cells. While it is excellent for visualizing cell morphology, it cannot be used alone to distinguish microglia from other myeloid populations.
Q: Can I use TMEM119 for the identification of human brain tissue?
A: Yes. TMEM119 is a reliable marker for microglia in both mouse and human brain tissue. It is highly specific to resident CNS myeloid cells and does not stain peripheral macrophages.
Q: Why is my P2RY12 staining weak in diseased tissue?
A: P2RY12 is significantly downregulated in activated and disease-associated microglia. This is a biological phenomenon, not a technical artifact. To accurately count all microglia in diseased tissue, use P2RY12 in combination with Iba1 or CD11b.
Q: How do I distinguish microglia from macrophages?
A: The most reliable approach is to use a combination of markers. Resident microglia are typically TMEM119+P2RY12+CD45lo, while infiltrating macrophages are TMEM119-P2RY12-CD45hi. However, note that activated microglia may upregulate CD45 and downregulate TMEM119 and P2RY12.
Q: What is the best marker for flow cytometry?
A: For flow cytometry, start by gating on CD45loCD11b+ cells to identify microglia. Then add TMEM119 or P2RY12 for further purification. For activation state analysis, use CD68, CD86, and CD206.
Q: Should I use multiple markers for identification?
A: Absolutely. No single marker is universally reliable for identifying microglia across all experimental conditions. Using multiple complementary markers significantly improves the accuracy of cell type identification.
Microglial markers are essential tools for understanding the role of microglia in CNS health and disease. The choice of marker depends on the experimental question, and researchers should use multiple markers to confirm cell identity and activation state. While significant progress has been made in identifying microglial markers, there are still many challenges, including a lack of specificity, activation state plasticity, and species differences.
By staying up-to-date with the latest research and using appropriate experimental approaches, researchers can accurately characterize microglial phenotypes and gain new insights into the mechanisms underlying neurological disorders. This knowledge will ultimately lead to the development of novel therapies that target microglia to treat CNS diseases.
References
[1] Vaughan DW, Peters A (1974). Neuroglial cells in the cerebral cortex of rats from young adult to old age: an electron microscopy study [J]. J Neurocytol. 3, 405–429.
[2] Banati R (2003). Neuropathological imaging: in vivo detection of glial activation as a measure of disease and adaptive change in the brain [J]. Brit Med Bull. 65, 121–131.
[3] Gomez Perdiguero E, Schulz C, Geissmann F (2013). Development and homeostasis of "resident" myeloid cells: the case of the microglia [J]. Glia. 61, 112–120.
[4] Kreutzberg GW (1996). Microglia: a sensor for pathological events in the CNS [J]. Trends Neurosci. 19, 312–318.
[5] Stence N, Waite M, Dailey E (2001). Dynamics of microglial activation: a confocal time-lapse analysis in hippocampal slices [J]. Glia. 33, 256–266.
[6] Giordana MT, Attanasio A, et al. (1994). Reactive cell proliferation and microglia following injury to the rat brain [J]. Neuropathol Appl Neurobiol. 20, 163–174.
[7] Dihne M, Block F, Korr H, Topper R (2001). Time course of glial proliferation and glial apoptosis following excitotoxic CNS injury [J]. Brain Res. 902, 178–189.
[8] Eugenin EA, Eckardt D, et al. (2001). Microglia at brain stab wounds express connexin 43 and in vitro form functional gap junctions after treatment with interferon gamma and tumour necrosis factor alpha [J]. Proc Natl Acad Sci USA. 98, 4190–4195.
[9] Jurga AM, Paleczna M, Kuter KZ (2020). Overview of General and Discriminating Markers of Differential Microglia Phenotypes [J]. Front Cell Neurosci. 14:198.
[10] Wang J, He W, Zhang J (2023). A richer and more diverse future for microglia phenotypes [J]. Heliyon. 9(4):e14713.
[11] McGeer PL, McGeer EG (1995). The inflammatory response system of brain: implications for therapy of Alzheimer and other neurodegenerative diseases [J]. Brain Res Rev. 21, 195–218.
[12] McGeer PL, McGeer EG (1996). Anti-inflammatory drugs in the fight against Alzheimer's disease [J]. Ann N Y Acad Sci. 777, 213–220.
[13] Barger SW, Harmon AD (1997). Microglial activation by Alzheimer amyloid precursor protein and modulation by apolipoprotein E [J]. Nature. 388, 878–881.
[14] Diemel LT, Copelman CA, Cuzner ML (1998). Macrophages in CNS remyelination: friend or foe [J]? Neurochem Res. 23, 341–347.
[15] Lees GJ (1993). The possible contribution of microglia and macrophages to delayed neuronal death after ischaemia [J]. J Neurol Sci. 114, 119–122.
[16] Tikka TM, Koistinaho JE (2001). Minocycline provides neuroprotection against N-methyl-D-aspartate neurotoxicity by inhibiting microglia [J]. J Immunol. 166, 7527–7533.
[17] Yun SP, Kam TI, et al. (2018). Block of A1 astrocyte conversion by microglia is neuroprotective in models of Parkinson's disease [J]. Nat Med. 24, 931–938.
[18] Fiala M, Liu QN, et al. (2002). Cyclooxygenase-2-positive macrophages infiltrate the Alzheimer's disease brain and damage the blood-brain barrier [J]. Eur J Clin Invest. 32, 360–371.
[19] Munder M (2009). Arginase: An emerging key player in the mammalian immune system [J]. Br J Pharmacol. 158(3), 638–651.
[20] Fabriek BO, et al. (2005). CD163-positive macrophages in inflammatory multiple sclerosis lesions: Distribution and impact on leukocyte recruitment [J]. J Neuroimmunol. 164(1–2), 106–114.
[21] Garden GA, Möller T. Microglia biology in health and disease [J]. J Neuroimmune Pharmacol. 2006 Jun;1(2):127-37.
[22] Stevens B, Allen NJ, et al. The classical complement cascade mediates CNS synapse elimination [J]. Cell. 2007 Dec 14;131(6):1164-78.
[23] Butovsky O, Jedrychowski MP, et al. Identification of a unique TGF-β-dependent molecular and functional signature in microglia [J] Nat Neurosci. 2014 Jan;17(1):131-43.
[24] Bennett ML, Bennett FC, et al. New tools for studying microglia in the mouse and human CNS [J]. Proc Natl Acad Sci U S A. 2016 Mar 22;113(12): E1738-46.
[25] Keren-Shaul H, Spinrad A, et al. A Unique Microglia Type Associated with Restricting Development of Alzheimer's Disease [J]. Cell. 2017 Jun 15;169(7):1276-1290.e17.
[26] Zhang Y, Chen K, et al. An RNA-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex [J]. J Neurosci. 2014 Sep 3;34(36):11929-47.


Comments
Leave a Comment