Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
C-C motif chemokine 17 (also known as TARC) is a chemokine belonging to the CC subfamily, primarily involved in regulating immune cell migration, activation, and inflammatory responses. It is produced by dendritic cells, endothelial cells, keratinocytes, and fibroblasts, and is highly expressed in the thymus, playing a critical role in T-cell development, migration, and activation of mature T cells. CCL22 and CCL17 both belong to the CC chemokine family. They share high homology, possess a common receptor molecule CCR4, and can effectively chemotax CCR4-expressing cells.
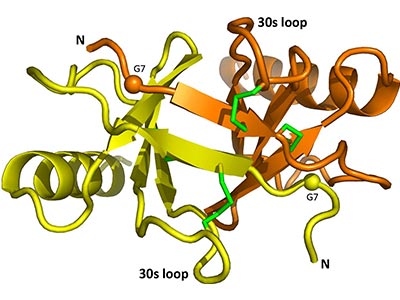
Figure 1. Schematic diagram of CCL17 dimer from the structure 1NR4. Spheres: G7T mutations, Green sticks: Disulfides.
Source: DOI: 10.1016/j.bbrep.2017.11.005
CCL17 plays a key role in cardiovascular diseases, particularly in age-related and angiotensin II (Ang II)-induced pathological cardiac remodeling and heart failure. Studies have found that serum CCL17 levels increase significantly with age and are associated with cardiac dysfunction. In animal experiments, CCL17 knockout mice exhibited inhibition of age- and Ang II-induced cardiac hypertrophy and fibrosis, accompanied by plasticity and differentiation of T-cell subsets [1]. Additionally, treatment with anti-CCL17 neutralizing antibodies significantly suppressed Ang II-induced pathological cardiac remodeling [2]. These findings highlight the potential of CCL17 as a novel therapeutic target for age-related and Ang II-induced pathological cardiac hypertrophy and heart failure.
The role of CCL17 in inflammation is primarily manifested through its binding to the CCR4 receptor, participating in the regulation of inflammatory responses and inflammatory pain. In inflammatory models, CCL17 promotes the chemotaxis and activation of inflammatory T cells, particularly in Th2-type immune responses [3]. In an osteoarthritis model, CCL17 knockout mice showed resistance to pain and disease progression, indicating a critical role of CCL17 in the pathogenesis of inflammatory pain [4]. Furthermore, in models of inflammatory arthritis and intestinal inflammation, the absence of CCL17 alleviated disease symptoms, further supporting its importance in inflammation [5][6].
In autoimmune diseases, CCL17 also plays a significant role [3]. Increased expression of CCL17 has been observed in systemic lupus erythematosus (SLE), suggesting its involvement in the immunopathological processes of SLE [7][8]. Additionally, in the experimental autoimmune encephalomyelitis (EAE) model of multiple sclerosis (MS), CCL17 knockout mice exhibited reduced disease symptoms [9]. These results indicate that CCL17 is crucial in the pathogenesis of autoimmune diseases and may serve as a new therapeutic target for these conditions.
CCL17 plays a key role in allergic asthma. Produced by dendritic cells (DCs), it attracts Th2 cells to the airways, triggering inflammatory responses in allergic asthma. NOD1-stimulated dendritic cells exacerbate Th2 pulmonary responses in vivo, and this effect occurs in a CCL17-dependent manner [10]. Studies using humanized SCID mouse models have shown that blocking CCR4 (the receptor for CCL17) significantly reduces airway inflammation and bronchial hyperresponsiveness, indicating that the CCR4-CCL17 axis is critical for recruiting Th2 cells to the airways [11]. These findings suggest that CCL17 may be a potential therapeutic target for allergic asthma.
CCL17 is highly expressed in various tumor cells and promotes tumor cell proliferation, migration, and invasion. In hepatocellular carcinoma, co-culture with M2-type macrophages or treatment with CCL17 enhances these malignant biological behaviors of tumor cells, improves cell stemness, promotes tumor sphere formation, and upregulates the expression of tumor stem cell-related transcription factors, facilitating tumor recurrence and metastasis [12]. Additionally, CCL17 influences the epithelial-mesenchymal transition process by activating the TGF-β1/Smad and Wnt/β-catenin signaling pathways, further enhancing the invasive and metastatic capabilities of tumor cells [13].
CCL17 is associated with the infiltration of multiple immune cells. In gastric and thyroid cancers, its high expression is linked to the accumulation of Foxp3+ regulatory T cells, potentially inhibiting anti-tumor immunity [13][14]. Meanwhile, the CCR4-CCL17 signaling axis selectively recruits Th2 cells, Tregs, and memory T cells to the inflammatory or tumor microenvironment, activating immunosuppressive programs through the JAK/STAT6 and PI3K/AKT pathways.
| Signal Pathway/Molecule | Description | Related Diseases/Cell Types | Reference |
|---|---|---|---|
| CCL17 / CCR4 | Promotes the migration, invasion, and stemness maintenance of tumor cells, and regulates the migration of immune cells | Multiple tumors / Stem cells | 12, 15, 16, 17, 18 |
| ERK / PD-L1 | CCL17 activates the ERK/PD - L1 signaling pathway through CCR4, enhancing the malignant behaviors of tumor cells | Esophageal squamous cell carcinoma | 15 |
| mTORC1 / mTORC2 | Lactic acid induces M2 polarization, M2 secretes CCL17, and CCL17/CCR4 activates mTORC1 to promote tumor invasion | Pituitary adenoma | 16, 19 |
| Wnt/β-catenin; TGF-β1 | CCL17 promotes EMT and stemness in tumor cells and activates Wnt/β-catenin and TGF-β1 signals | Hepatocellular carcinoma | 12 |
| STAT6 / IRF4/JMJD3 | The expression of CCL17 induced by IL-4 depends on the STAT6-IRF4-JMJD3 pathway and involves epigenetic regulation | Monocytes / Macrophages | 7 |
| β-arrestin | CCL17 activates CCR4 mainly by recruiting β-arrestin rather than G protein, suggesting that it may act as a scavenger receptor | T cells | 20 |
High expression of CCL17/CCL22 can predict the efficacy of immune checkpoint inhibitors and survival rate in patients with head and neck squamous cell carcinoma [19], and also has potential diagnostic and prognostic value in diseases such as nasal NK/ T-cell lymphoma and liver cancer [12][18].
Anti-CCR4 monoclonal antibodies can mediate NK cells to kill tumor cells, and the CCL17/CCR4 axis has become a novel targeted therapy candidate for various tumors [18][21].
To support the research on CCL17 by pharmaceutical companies and research institutions, CUSABIO has launched CCL17 recombinant protein, CCL17 recombinant antibody and ELISA kits to assist your research on the mechanisms of CCL17 or its potential clinical value.
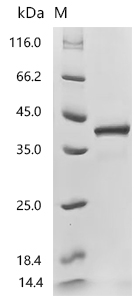
● CCL17 Recombinant Protein
Recombinant Human C-C motif chemokine 17 (CCL17) (Active); CSB-MP856406HU
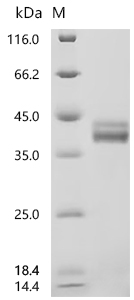
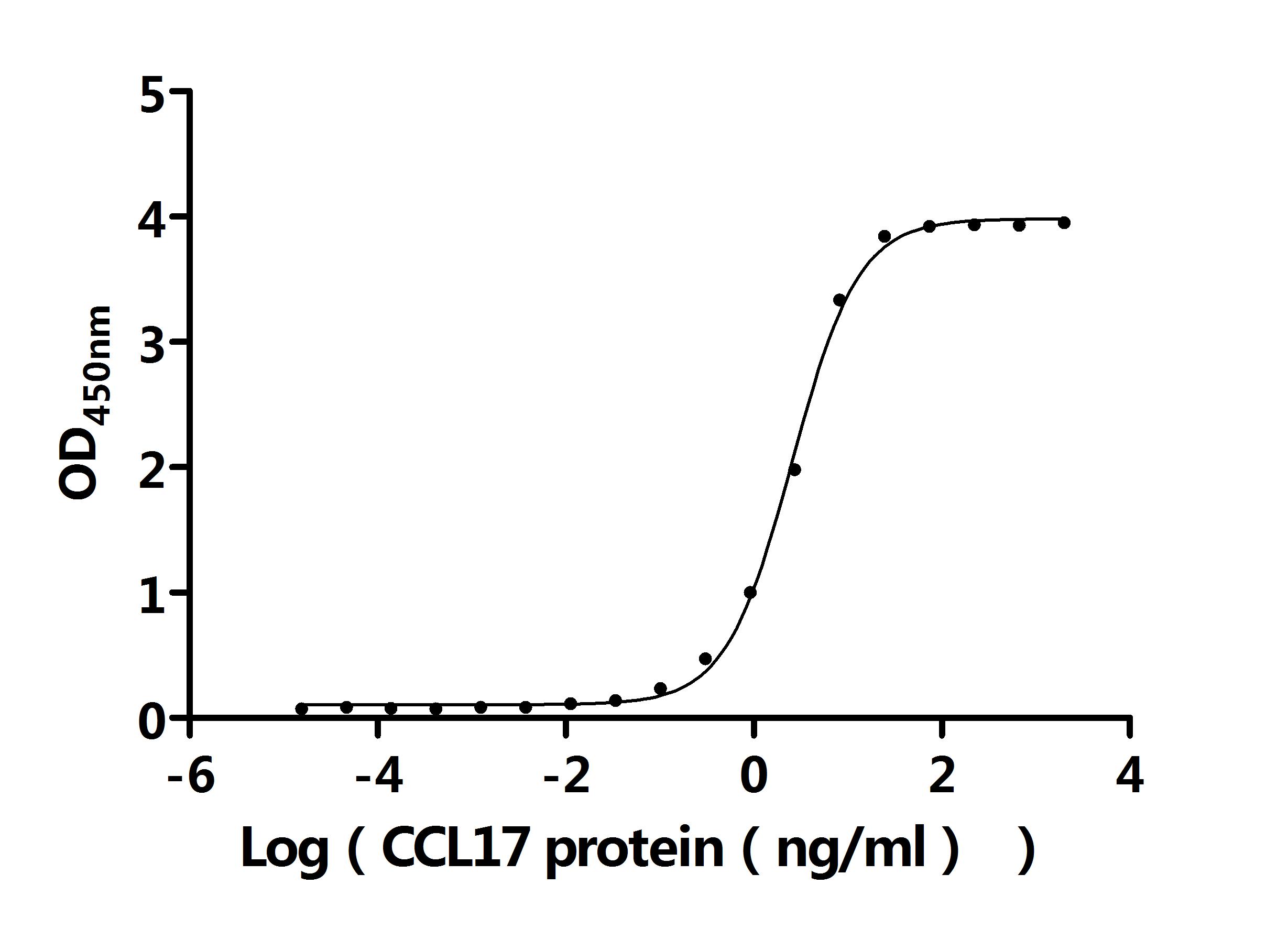
Recombinant Macaca mulatta C-C motif chemokine 17 (CCL17) (Active); CSB-MP811562MOW
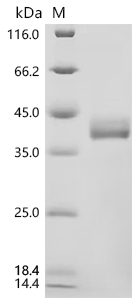
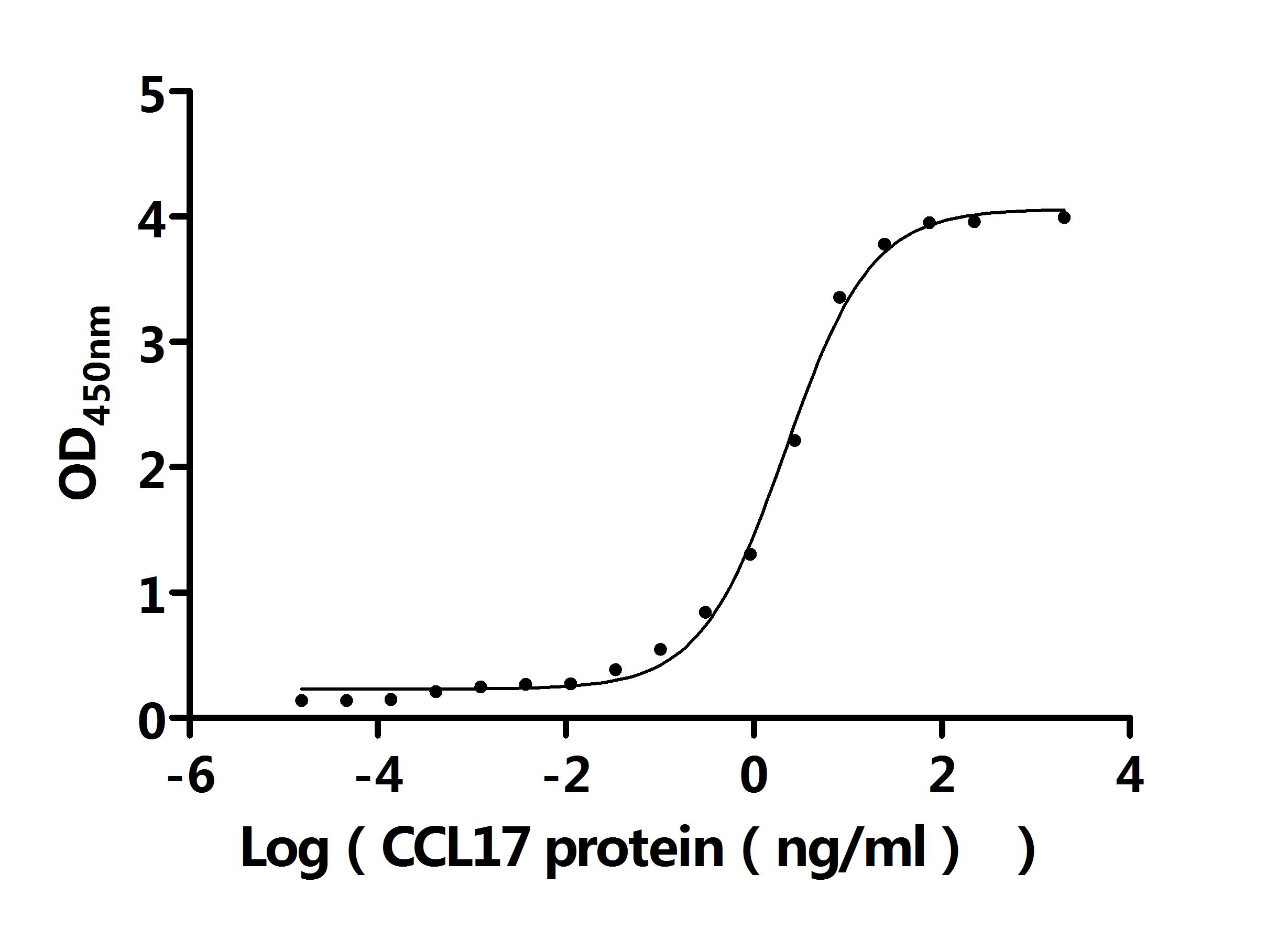
Recombinant Mouse C-C motif chemokine 17 (Ccl17) (Active); CSB-MP6512MO
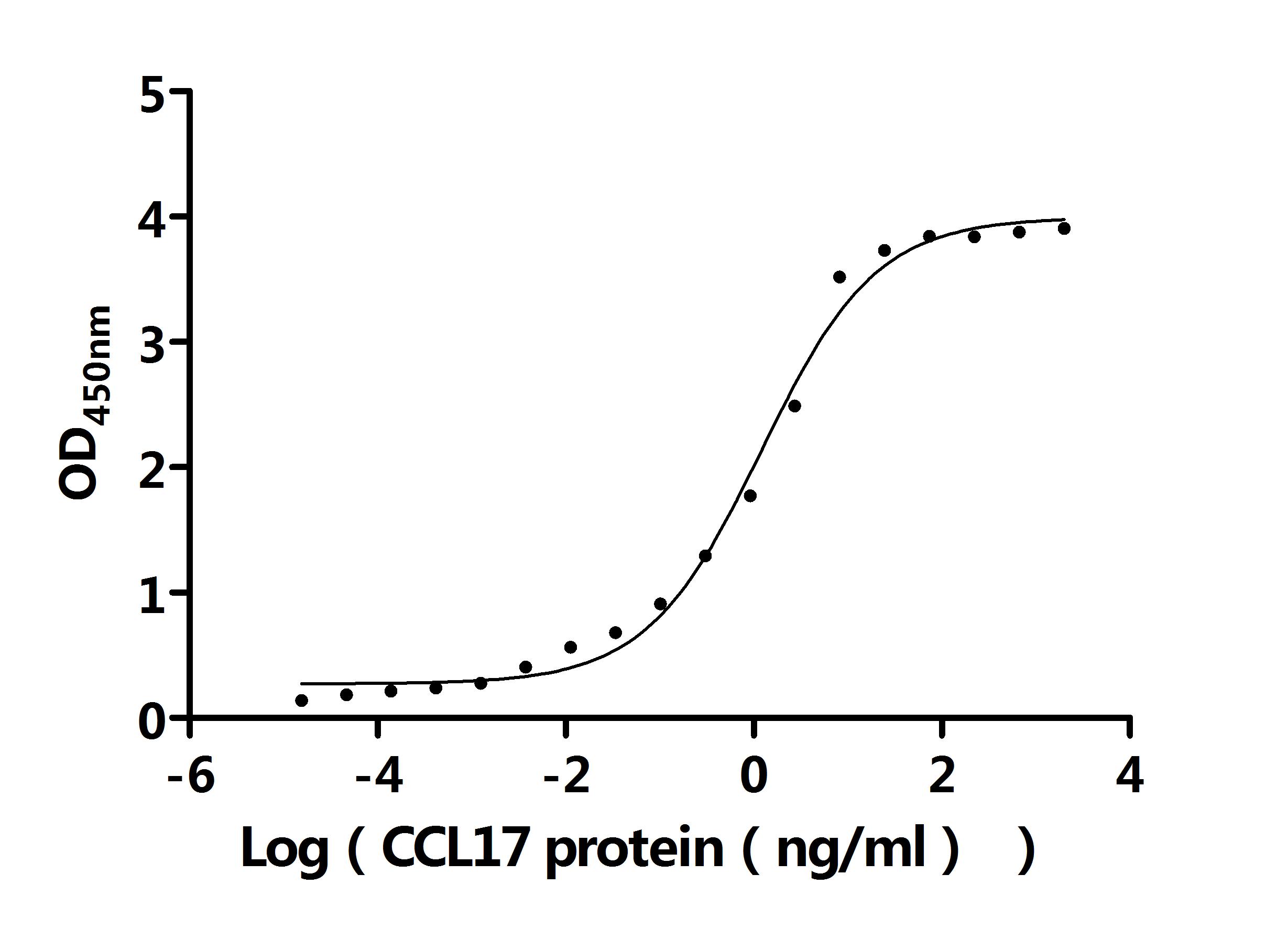
● CCL17 Recombinant Antibody
CCL17 Recombinant Monoclonal Antibody; CSB-RA856406MA1HU
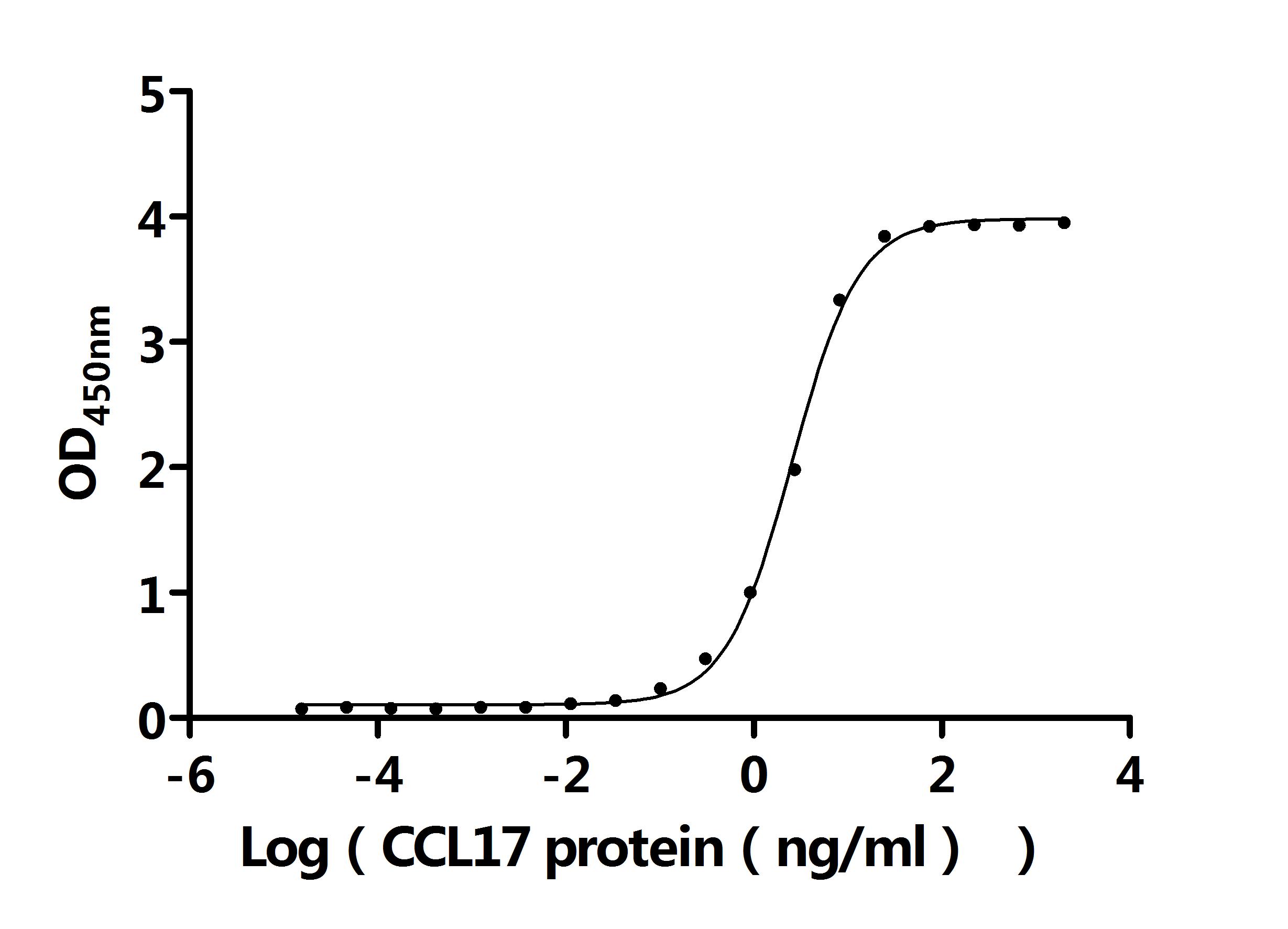
Activity: Measured by its binding ability in a functional ELISA. Immobilized Anti-CCL17 recombinant antibody at 2 μg/mL can bind Human CCL17 protein (CSB-MP856406HU). The EC50 is 2.403-2.741 ng/mL.
References
[1] Zhang, Y., Ye, Y., Tang, X., Wang, H., Tanaka, T., Tian, R., Yang, X., Wang, L., Xiao, Y., Hu, X., Jin, Y., Pang, H., Du, T., Liu, H., Sun, L., Xiao, S., Dong, R., Ferrucci, L., Tian, Z., & Zhang, S. (2022). CCL17 acts as a novel therapeutic target in pathological cardiac hypertrophy and heart failure. The Journal of Experimental Medicine, 219(8).
[2] Zhang, Y., Tang, X., Wang, Z., Wang, L., Chen, Z., Qian, J. Y., Tian, Z., & Zhang, S. Y. (2023). The chemokine CCL17 is a novel therapeutic target for cardiovascular aging. Signal transduction and targeted therapy, 8(1), 157.
[3] Lupancu, T.J., Eivazitork, M., Hamilton, J.A., Achuthan, A.A. and Lee, K.M.-C. (2023), CCL17/TARC in autoimmunity and inflammation—not just a T-cell chemokine. Immunol Cell Biol, 101: 600-609.
[4] Lee, M.-C., Saleh, R., Achuthan, A., Fleetwood, A. J., Förster, I., Hamilton, J. A., & Cook, A. D. (2018). CCL17 blockade as a therapy for osteoarthritis pain and disease. Arthritis Research & Therapy, 20(1), 62.
[5] Heiseke, A. F., Faul, A. C., Lehr, H., Förster, I., Schmid, R. M., Krug, A. B., & Reindl, W. (2011). CCL17 Promotes Intestinal Inflammation in Mice and Counteracts Regulatory T Cell–Mediated Protection From Colitis. Gastroenterology, 142(2), 335–345.
[6] Achuthan, A., Cook, A. D., Kevin M.-C. Lee, Saleh, R., Hsu Wei Khiew, Melody Wei-Ning Chang, Louis, C., Fleetwood, A. J., Lacey, D., Anne Deen Christensen, Frye, A. T., Pui Yeng Lam, Kusano, H., Nomura, K., Steiner, N., Förster, I., Nutt, S. L., Olshansky, M., Turner, S. J., & Hamilton, J. A. (2016). Granulocyte macrophage colony-stimulating factor induces CCL17 production via IRF4 to mediate inflammation. Journal of Clinical Investigation, 126(9), 3453–3466.
[7] Hsu, A. T., Lupancu, T. J., Lee, M.-C., Fleetwood, A. J., Cook, A. D., Hamilton, J. A., & Achuthan, A. (2018). Epigenetic and transcriptional regulation of IL4-induced CCL17 production in human monocytes and murine macrophages. Journal of Biological Chemistry, 293(29), 11415–11423.
[8] T R D J Radstake. (2004). Increased expression of CCL18, CCL19, and CCL17 by dendritic cells from patients with rheumatoid arthritis, and regulation by Fc gamma receptors. Annals of the Rheumatic Diseases, 64(3), 359–367.
[9] Dhaiban, S., Al-Ani, M., Elemam, N. M., & Maghazachi, A. A. (2020). Targeting Chemokines and Chemokine Receptors in Multiple Sclerosis and Experimental Autoimmune Encephalomyelitis. Journal of Inflammation Research, Volume 13, 619–633.
[10] Yahia, S. A., Imane Azzaoui, Laetitia Everaere, Vorng, H., Cécile Chenivesse, Philippe Marquillies, Duez, C., Delacre, M., Grandjean, T., Balsamelli, J., d’Andon, M. F., Fan, Y., Coline Ple, Werts, C., Ivo Gomperts Boneca, Wallaert, B., Mathias Chamaillard, & Tsicopoulos, A. (2014). CCL17 Production by Dendritic Cells Is Required for NOD1-mediated Exacerbation of Allergic Asthma. American Journal of Respiratory and Critical Care Medicine, 189(8), 899–908.
[11] Perros, F., Hoogsteden, H. C., Coyle, A. J., Lambrecht, B. N., & Hammad, H. (2009). Blockade of CCR4 in a humanized model of asthma reveals a critical role for DC-derived CCL17 and CCL22 in attracting Th2 cells and inducing airway inflammation. Allergy, 64(7), 995–1002.
[12] Zhu, F., Li, X., Chen, S., Zeng, Q., Zhao, Y., & Luo, F. (2016). Tumor-associated macrophage or chemokine ligand CCL17 positively regulates the tumorigenesis of hepatocellular carcinoma. Medical Oncology, 33(2).
[13] Mizukami, Y., Kono, K., Kawaguchi, Y., Akaike, H., Kamimura, K., Sugai, H., & Fujii, H. (2008). CCL17 and CCL22 chemokines within tumor microenvironment are related to accumulation of Foxp3+ regulatory T cells in gastric cancer. International Journal of Cancer, 122(10), 2286–2293.
[14] Gu, X., Chen, B., Zhang, S., Zhai, X., Hu, Y., & Ye, H. (2024). The expression of CCL17 and potential prognostic value on tumor immunity in thyroid carcinoma based on bioinformatics analysis. Scientific Reports, 14(1).
[15] Jin, C., Lu, L., Gao, J., & Chen, L. (2024). M2-like Macrophages-derived CCL17 Promotes Esophageal Squamous Cell Carcinoma Metastasis and Stemness via Activating CCR4-mediated ERK/PD-L1 Pathway.. Current molecular medicine.
[16] Zhang, A., Xu, Y., Xu, H., Ren, J., Meng, T., Ni, Y., Zhu, Q., Zhang, W., Pan, Y., Jin, J., Bi, Y., Wu, Z., Lin, S., & Lou, M. (2021). Lactate-induced M2 polarization of tumor-associated macrophages promotes the invasion of pituitary adenoma by secreting CCL17. Theranostics, 11, 3839 - 3852.
[17] Konno, K., Sasaki, T., Kulkeaw, K., & Sugiyama, D. (2020). Paracrine CCL17 and CCL22 signaling regulates hematopoietic stem/progenitor cell migration and retention in mouse fetal liver.. Biochemical and biophysical research communications.
[18] Kumai, T., Nagato, T., Kobayashi, H., Komabayashi, Y., Ueda, S., Kishibe, K., Ohkuri, T., Takahara, M., Celis, E., & Harabuchi, Y. (2015). CCL17 and CCL22/CCR4 signaling is a strong candidate for novel targeted therapy against nasal natural killer/T-cell lymphoma. Cancer Immunology, Immunotherapy, 64, 697-705.
[19] Zhou, W., Zhang, X., Feng, Y., Zhang, Y., & Liu, Z. (2022). The CC ligand chemokine family members CCL17/CCL22 predict the survival and response to immune checkpoint blockade therapy of patients with head and neck squamous cell carcinoma.. Current problems in cancer, 46 6, 100896 .
[20] Lim, H., Lane, J., Canals, M., & Stone, M. (2021). Systematic Assessment of Chemokine Signaling at Chemokine Receptors CCR4, CCR7 and CCR10. International Journal of Molecular Sciences, 22.
[21] Goenka, A., Khan, F., Verma, B., Sinha, P., Dmello, C., Jogalekar, M., Gangadaran, P., & Ahn, B. (2023). Tumor microenvironment signaling and therapeutics in cancer progression. Cancer Communications, 43, 525 - 561.


Comments
Leave a Comment