Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
The CHRNA1 gene is located on human chromosome 2q31.1 and encodes the α1 subunit of the nicotinic acetylcholine receptor (nAChR). The protein consists of 457 amino acids and contains four transmembrane domains (M1–M4), with M2 forming the ion channel pore responsible for cation selectivity [1]. At the neuromuscular junction (NMJ), CHRNA1 assembles with β1, δ, and ε/γ subunits to form a pentameric receptor, mediating acetylcholine (ACh) signaling that induces muscle cell depolarization and contraction [2][3].
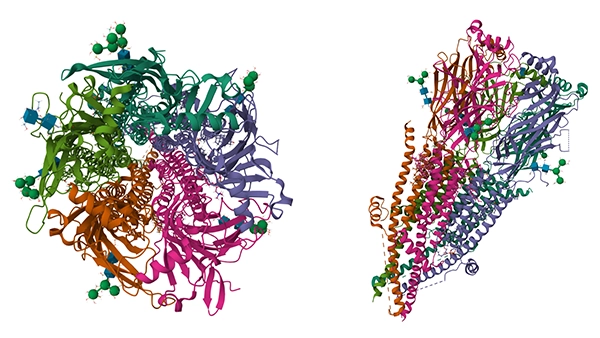
Human muscle nAChR apo state (from:PDB)
Knockout studies show that the absence of CHRNA1 causes abnormal NMJ development and completely blocks synaptic transmission, highlighting its essential role in NMJ formation and maintenance [2]. In addition, CHRNA1 interacts with the scaffolding protein rapsyn to ensure high-density receptor clustering on the postsynaptic membrane [4]. Phosphorylation sites within the M3–M4 intracellular loop regulate receptor internalization, thereby contributing to synaptic plasticity [3]. In adult skeletal muscle, the ε subunit replaces the fetal γ subunit, significantly increasing channel conductance; this subunit switch is tightly regulated by the agrin-MuSK signaling pathway [5].
CHRNA1 is highly expressed in NMJs of the peripheral nervous system, where it colocalizes with acetylcholinesterase to form efficient postsynaptic specializations [6][7]. Although expression levels in the central nervous system are lower, CHRNA1 has been detected in the hippocampus, cortex, and brainstem, suggesting roles in cognition and autonomic regulation [8].
In thymic medullary epithelial cells, ectopic expression of CHRNA1—regulated by AIRE—is associated with central immune tolerance and may trigger autoantibody production in myasthenia gravis [6]. Single-cell sequencing has shown stage-specific peaks of CHRNA1 expression in spinal motor neurons during NMJ maturation [9]. CHRNA1 transcripts are also detectable in pulmonary artery smooth muscle cells, with expression negatively correlated with mitochondrial complex I activity, implicating potential roles in vascular tone regulation [10].
As the core nAChR subunit, the extracellular N-terminal domain of CHRNA1 forms part of the ACh-binding site, while the M2 transmembrane helix lines the ion-conducting pore [5]. CHRNA1–β1 interactions regulate channel opening probability; β1 mutations reduce current amplitude by ~60% [11]. During development, γ-containing fetal-type receptors are replaced by ε-containing adult-type receptors, altering receptor kinetics [1].
CHRNA1 can also form heteromeric receptors with neuronal subunits such as α7 or α3β4, modifying calcium permeability and desensitization kinetics [12]. Moreover, CHRNA1 has been shown to interact with 5-HT3A and GABAA receptors, influencing current decay and receptor sensitivity [11][2]. In pathological states such as COPD, CHRNA1–α9 colocalization is lost, accompanied by increased inflammatory cytokine secretion [11].
Upon ACh release from motor neurons, two ACh molecules bind to CHRNA1, triggering conformational changes that open the ion channel, generating an endplate potential and activating voltage-gated sodium channels to initiate muscle contraction [5]. Postsynaptic clustering of CHRNA1 depends on the agrin-LRP4-MuSK-rapsyn pathway [5]. In MuSK knockout mice, CHRNA1 expression decreases by 80%, and postsynaptic folds are absent [5].
PKA-mediated phosphorylation enhances receptor sensitivity, while CaMKII accelerates desensitization to prevent excitotoxicity [13]. At the transcriptional level, neuregulin-1 and MuSK signaling promote CHRNA1 expression and subunit switching. Genetic polymorphisms such as rs16862847 are associated with reduced endplate potentials [12]. In COPD, downregulated CHRNA1 expression correlates with impaired cholinergic signaling [11].
CHRNA1 plays a pivotal role in the agrin-MuSK pathway, ensuring NMJ stability. Mutations in LRP4 disrupt MuSK phosphorylation and CHRNA1 localization, leading to congenital myasthenic syndromes (CMS) [3]. Anti-MuSK antibodies also impair CHRNA1 anchoring at the postsynaptic membrane [1]. Additionally, CHRNA1 expression may influence immune tolerance through mTOR signaling in thymic epithelial cells, indicating a role in neuro-immune crosstalk [5].
In thymic epithelial cells, CHRNA1 modulates immune tolerance via the mTOR pathway. CHRNA1 deficiency reduces mTOR activity, leading to thymic developmental defects and autoimmune responses [9][14]. Interaction between CHRNA1 and STAT3 activates the Akt/mTOR cascade, as demonstrated in nicotine-induced atherosclerosis models [15]. Dysregulated CHRNA1–mTOR signaling may contribute to autoimmune myasthenia gravis [7].
Mutations in CHRNA1 account for ~10–15% of CMS cases, mostly inherited in an autosomal recessive manner [1]. Missense and nonsense variants reduce receptor sensitivity or surface expression, significantly lowering endplate potentials [5]. Clinically, patients present in infancy with ptosis, feeding difficulties, and generalized weakness, and often show poor response—or even worsening—to pyridostigmine treatment [16]. Some variants, such as V285L, cause temperature-sensitive myasthenia due to protein instability [1].
CHRNA1 may contribute to PAH pathogenesis by regulating mitochondrial electron transport and reactive oxygen species (ROS) production. CHRNA1 is upregulated in PAH lung tissues, correlating with decreased ETC complex II–III activity [10]. HIF-1α can activate CHRNA1 transcription under hypoxia [11]. Knockdown of CHRNA1 alleviates metabolic reprogramming and apoptosis resistance in PASMCs [10]. In animal models, CHRNA1 antagonists reduce pulmonary arterial pressure, supporting its potential as a therapeutic target [10].
CHRNA1 mRNA is significantly elevated in OA synovium, positively correlated with IL-6 and TNF-α expression [2]. Multi-omics studies suggest that gut microbiota influence CHRNA1 expression through the "gut–muscle–joint axis," affecting joint homeostasis [17]. Abnormal CHRNA1 expression is associated with impaired autophagy, mitochondrial dysfunction, and cartilage degeneration [12][17]. Genetic polymorphisms are linked to OA pain sensitivity and progression [8].
Recent studies have explored several therapeutic strategies targeting CHRNA1. For primary focal hyperhidrosis, experimental evidence indicates that PAI1 may suppress CHRNA1 expression, thereby reducing sweat gland secretion, whereas inhibition of PAI1 with PAI-039 or PAI1 gene knockout enhances CHRNA1-related hyperhidrotic phenotypes. In contrast, the CHRNA1 antagonist cisatracurium alleviates hyperhidrosis by blocking receptor ion channels.
For congenital myasthenic syndromes (CMS) caused by CHRNA1 mutations, antisense oligonucleotides (AONs) targeting the exon P3A 5′ splice site have shown the potential to restore normal splicing balance, representing a promising therapeutic approach. Moreover, CHRNA1 genetic polymorphisms may influence patient sensitivity and adverse reaction risks to neuromuscular blockers such as rocuronium, providing a genetic basis for individualized therapy.
CHRNA1, encoding the α1 subunit of nAChR, is essential for NMJ signaling and is implicated in MuSK and mTOR pathways. Its mutations or abnormal expression are closely associated with CMS, PAH, and OA.
CUSABIO offers CHRNA1-related proteins and antibodies as research resources to facilitate studies on molecular mechanisms and therapeutic targeting, supporting both basic and translational research.
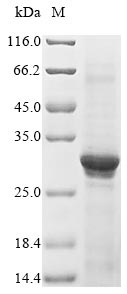
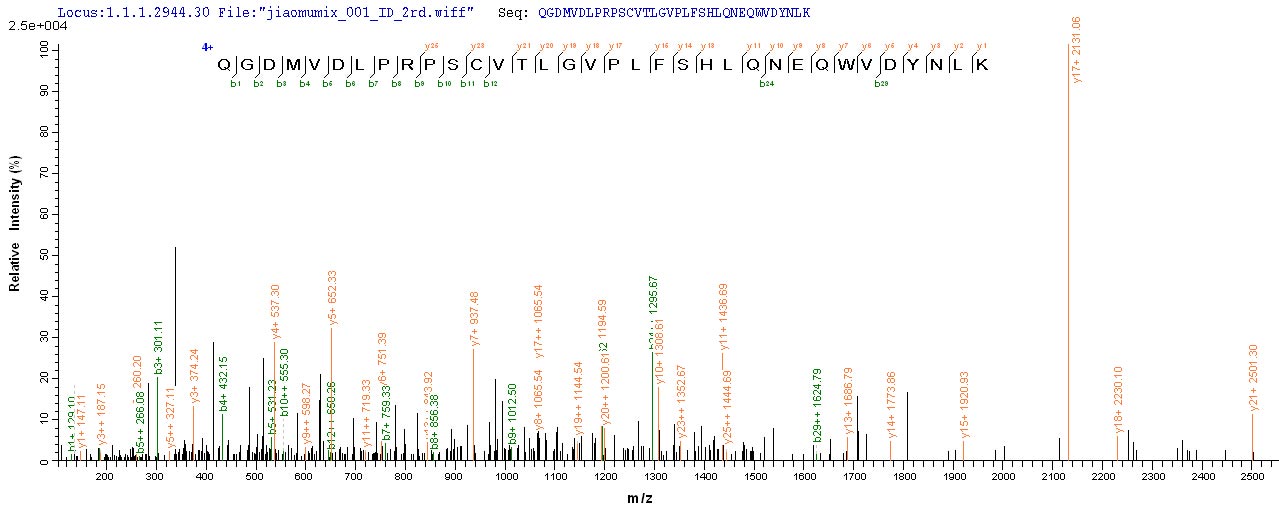
● CHRNA1 Recombinant Protein
Recombinant Human Acetylcholine receptor subunit alpha (CHRNA1), partial; CSB-EP005386HU
References
[1] Pedro M. Rodríguez Cruz, Jacqueline Palace, David Beeson. Congenital myasthenic syndromes and the neuromuscular junction[J]. Current Opinion in Neurology, 2014, 27(5): 566-575.
[2] Qin Chen, Shaosheng Bei, Zhiyun Zhang, Xiaofeng Wang, Yunying Zhu. Identification of diagnostic biomarks and immune cell infiltration in ulcerative colitis[J]. Scientific Reports, 2023, 13(1).
[3] Bisei Ohkawara, Macarena Cabrera‐Serrano, Tomohiko Nakata, Margherita Milone, Nobuyuki Asai, Kenyu Ito, Mikako Ito, Akio Masuda, Yasutomo Ito, Andrew G. Engel, Kinji Ohno. LRP4 third β-propeller domain mutations cause novel congenital myasthenia by compromising agrin-mediated MuSK signaling in a position-specific manner[J]. Human Molecular Genetics, 2013, 23(7): 1856-1868.
[4] Kai Qian, Jiaxin Xu, Yi Deng, Hao Peng, Jun Peng, Chun‐Mei Ou, Liu Zu, Lihong Jiang, Yonghang Tai. Signaling pathways of genetic variants and miRNAs in the pathogenesis of myasthenia gravis[J]. Gland Surgery, 2020, 9(6): 1933-1944.
[5] Bisei Ohkawara, Mikako Ito, Kinji Ohno. Secreted Signaling Molecules at the Neuromuscular Junction in Physiology and Pathology[J]. International Journal of Molecular Sciences, 2021, 22(5): 2455-2455.
[6] Matthieu Giraud, Richard Taubert, Claire Vandiedonck, Xiayi Ke, Matthieu Lévi‐Strauss, Franco Pagani, Francisco E. Baralle, B. Eymard, Christine Tranchant, Philippe Gajdos, Angela Vincent, Nick Willcox, David Beeson, Bruno Kyewski, Henri-Jean Garchon. An IRF8-binding promoter variant and AIRE control CHRNA1 promiscuous expression in thymus[J]. Nature, 2007, 448(7156): 934-937.
[7] Matthieu Giraud, Claire Vandiedonck, Henri‐Jean Garchon. Genetic Factors in Autoimmune Myasthenia Gravis[J]. Annals of the New York Academy of Sciences, 2008, 1132(1): 180-192.
[8] Peter Mu‐Hsin Chang, Yi‐Chen Yeh, Tzu-Chi Chen, Yu–Chung Wu, Pei‐Jung Lu, Hui-Chuan Cheng, Hsueh‐Ju Lu, Ming‐Huang Chen, Teh‐Ying Chou, Chi‐Ying F. Huang. High Expression of CHRNA1 is Associated with Reduced Survival in Early Stage Lung Adenocarcinoma after Complete Resection[J]. Annals of Surgical Oncology, 2013, 20(11): 3648-3654.
[9] Zhanfeng Liang, Lianjun Zhang, Huiting Su, Rong Luan, Ning Na, Lina Sun, Yang Zhao, Xiaodong Zhang, Qian Zhang, Juan Li, Lianfeng Zhang, Yong Zhao. MTOR signaling is essential for the development of thymic epithelial cells and the induction of central immune tolerance[J]. Autophagy, 2017, 14(3): 505-517.
[10] Xin Zhang, Jieling Li, Minyi Fu, Xijie Geng, Junjie Hu, Kejing Tang, Pan Chen, Jianyong Zou, Xiaoman Liu, Bo Zeng. Dysfunction in mitochondrial electron transport chain drives the pathogenesis of pulmonary arterial hypertension: insights from a multi-omics investigation[J]. Respiratory Research, 2025, 26(1).
[11] Lin Chen, Donglan Zhu, Jinfu Huang, Hui Zhang, Guang Zhou, Xiaoning Zhong. Identification of Hub Genes Associated with COPD Through Integrated Bioinformatics Analysis[J]. International Journal of COPD, 2022, Volume 17: 439-456.
[12] Patrick F. McArdle, Sue Rutherford, Braxton D. Mitchell, Coleen Damcott, Ying Wang, Ramachandran S. Vasan, Sandy Ott, Yen‐Pei C. Chang, Daniel Levy, Nanette Steinle. Nicotinic acetylcholine receptor subunit variants are associated with blood pressure
[13] Peter V. Lovell, David F. Clayton, Kirstin Replogle, Claudio V. Mello. Birdsong "Transcriptomics": Neurochemical Specializations of the Oscine Song System[J]. PLoS ONE, 2008, 3(10): e3440-e3440.
[14] Fatemeh Shirafkan, Luca Hensel, Kristin Rattay. Immune tolerance and the prevention of autoimmune diseases essentially depend on thymic tissue homeostasis[J]. Frontiers in Immunology, 2024, 15.
[15] Shuang Xu, Huaner Ni, Hangwei Chen, Qiuyan Dai. The interaction between STAT3 and nAChRα1 interferes with nicotine-induced atherosclerosis via Akt/mTOR signaling cascade[J]. Aging, 2019, 11(19): 8120-8138.
[16] Marco Calabrò, Laura Mandelli, Concetta Crisafulli, Antonella Sidoti, Tae‐Youn Jun, Soo-Jung Lee, Hans‐Jürgen Möller, Ashwin A. Patkar, Prakash S. Masand, Chi‐Un Pae, Alessandro Serretti. Genes Involved in Neurodevelopment, Neuroplasticity, and Bipolar Disorder: CACNA1C, CHRNA1, and MAPK1[J]. Neuropsychobiology, 2016, 74(3): 159-168.
[17] Tianyang Xu, Dong Kwon Yang, Kaiyuan Liu, Qiuming Gao, Zhongchen Liu, Guodong Li. Miya Improves Osteoarthritis Characteristics via the Gut-Muscle-Joint Axis According to Multi-Omics Analyses[J]. Frontiers in Pharmacology, 2022, 13.


Comments
Leave a Comment