

Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
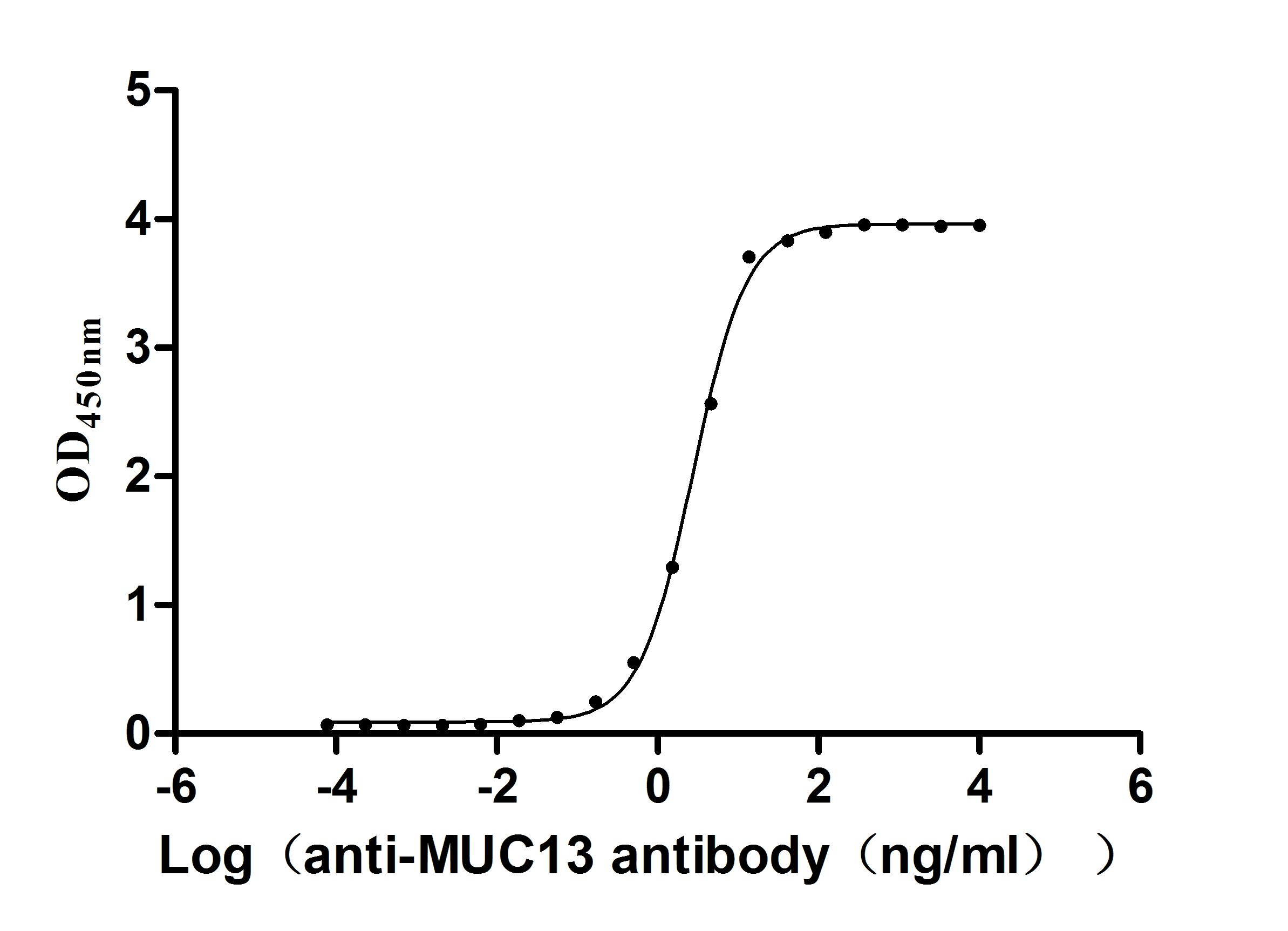
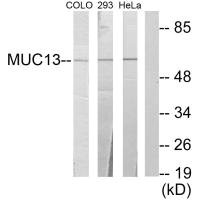
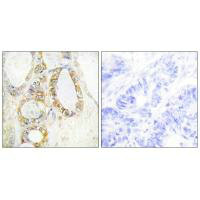
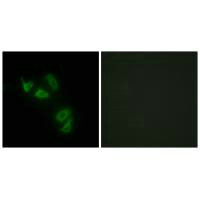
Transmembrane mucin MUC13(Mucin 13)It is an important component of the epithelial barrier and a signaling regulatory protein of great interest in tumor biology. Its structure includes a highly glycosylated extracellular domain and a cytoplasmic tail containing multiple phosphorylation sites, serving dual functions in barrier maintenance and signal integration. In digestive system tumors such as colorectal cancer, pancreatic cancer, and gastric cancer, as well as non-digestive system malignancies like lung cancer and ovarian cancer, MUC13 exhibits significant overexpression and loss of polarity, and throughBy activating key pathways such as NF-κB, YAP1/β-catenin, and HER2/ERK/Akt, it promotes tumorigenesis, invasion and metastasis, metabolic reprogramming, and chemotherapy resistance. This article systematically reviews the molecular structure, genetic and post-translational modification characteristics of MUC13, analyzes its pathogenic mechanisms in multi-system tumors, summarizes the progress in targeted drug development, and evaluates its translational potential as a broad-spectrum biomarker and therapeutic target.
1. Discovery, Structural Classification, and Tissue Distribution Characteristics of MUC13
2. Genetic abnormalities and post-translational modification characteristics of MUC13.
3. MUC13-mediated key signal transduction network
4. Pathogenic Mechanisms of MUC13 in Multisystem Malignancies
MUC13 belongs to the membrane-bound mucin family and is an important structural protein for maintaining epithelial integrity. This gene was initially identified by screening serine/threonine-rich sequences in the GenBank EST database and is located in the chromosomal region 3q13.3 [1]. Its full-length cDNA is approximately 3.2 kb, encoding a transmembrane glycoprotein of 512 amino acids.
At the structural level, MUC13 exhibits a typical yet complex multi-domain configuration: the N-terminus consists of a highly glycosylated tandem repeat region, followed by three EGF-like domains, a SEA module, a single transmembrane region, and a cytoplasmic tail containing multiple phosphorylatable sites [2,4]. This structural partitioning endows MUC13 with dual functions: the extracellular domain forms a physical barrier and participates in intercellular interface regulation, while the intracellular tail serves as a signaling integration node involved in cell fate regulation.
Under normal physiological conditions, MUC13 is primarily expressed in the colon, small intestine, and tracheal epithelium, with moderate expression observed in the kidneys, stomach, and appendix [1]. Its spatial localization is highly polarized, predominantly distributed in the apical membrane regions of columnar cells and goblet cells. This polarized expression is crucial for maintaining epithelial barrier homeostasis.
However, in the context of inflammation or malignant transformation, MUC13 expression levels are significantly elevated, accompanied by a shift in subcellular localization, including cytoplasmic and nuclear distribution [14,16]. This spatial remodeling is often accompanied by disorganization of tissue structure, serving as a significant hallmark of tumor progression.
MUC13 exhibits significant upregulation of mRNA in various epithelial tumors, and its enhanced expression is often associated with an increase in gene copy number [5]. Unlike certain mucins that frequently harbor mutations, the pathogenicity of MUC13 primarily manifests through expression reprogramming and ectopic expression, rather than specific driver mutations.
Post-translational modifications, particularly aberrant O-glycosylation, are key mechanisms determining its oncogenic function. GalNAc-T14-mediated glycosylation enhancement can alter the conformation and stability of MUC13 and augment its migration-related functions [6]. This process is regulated by the ERK1/2 signaling pathway, reflecting the coupling between glycosylation and intracellular signaling networks.
Furthermore, the cytoplasmic tail of MUC13 is rich in serine and tyrosine residues, providing a structural basis for its involvement in downstream kinase cascade reactions [4]. This functional transformation from a "structural protein" to a "signaling regulatory molecule" serves as the molecular prerequisite for its critical role in cancer.
In pancreatic and colorectal cancers, MUC13 regulates cellular metabolism and anti-apoptotic capacity by activating the NF-κB pathway. It induces nuclear translocation of p65 and phosphorylation of IκB, while upregulating GLUT-1 expression [7]. Beyond transcriptional regulation, MUC13 physically interacts with GLUT-1, enhancing glucose uptake and lactate production, thereby promoting the shift toward aerobic glycolysis.
In colorectal cancer, MUC13 enhances the stability of the TNFR1 complex by recruiting cIAP1 and RIPK1, thereby strengthening NF-κB signaling [8]. Simultaneously, in the context of DNA damage, it regulates the activation of the anti-apoptotic protein BCL-XL expression through ATM and NEMO, enhancing chemotherapy tolerance [8].
Furthermore, MUC13 regulates the distribution of tight junction proteins through PKC signaling, negatively modulating the integrity of the intestinal epithelial barrier [9]. This mechanism highlights its role in microenvironment remodeling.
MUC13 enhances resistance to anoikis during metastasis by constructing the YAP1/β-catenin nuclear complex [11,12]. RUNX1 can upregulate MUC13 expression and activate the Wnt/β-catenin signaling pathway [10].
In the non-anchored state, MUC13 promotes the nuclear translocation of YAP1 and synergistically upregulates c-Myc and Axin2 with β-catenin [13]. Animal models have confirmed that high expression of MUC13 significantly enhances distal colonization ability [12]. Knockdown of MUC13 markedly reduces spheroid formation capacity and the number of metastatic foci [13].
The abnormal expression of MUC13 is not confined to a single organ; rather, it exhibits highly consistent features of upregulated expression, aberrant localization, and signaling pathway remodeling across various epithelial-derived malignant tumors. Whether in digestive or non-digestive system tumors, its core pathogenic mechanism revolves around the continuous process of "enhanced expression—spatial reorganization—signal amplification—invasion and metastasis—drug resistance formation."
In colorectal cancer, the expression level of MUC13 shows a continuous increase from normal mucosa to primary tumors and further to metastatic lesions [14,15]. In addition to increased expression, its subcellular localization also undergoes significant changes, shifting from the original apical membrane localization to cytoplasmic and nuclear distribution [16]. This localization shift is significantly associated with low differentiation and poor prognosis. STAT5B can directly bind to its promoter region to promote transcriptional activation [16], while high expression of MUC13 further upregulates proliferation-related molecules such as Shh, Bmi-1, and TERT [16], and enhances extracellular matrix degradation capacity by regulating MMP1 [15].
During the metastatic phase, MUC13 enhances the nuclear aggregation of YAP1 and β-catenin to form a survival complex, which activates Bcl-2 and inhibits Caspase-3 cleavage, thereby conferring resistance to anoikis in tumor cells [17]. This mechanism directly explains its role in promoting distal colonization. The downregulation of miR-4647 leads to uncontrolled MUC13 expression [18], while the circRNA regulatory network also participates in shaping the microenvironment [19]. Additionally, MUC13 enriched in exosomes provides a potential molecular marker basis for liquid biopsy [20].
In pancreatic ductal adenocarcinoma (PDAC), MUC13 is also significantly overexpressed [21]. Multi-omics analysis reveals that the long transcript L-MUC13 exhibits stronger tumorigenic potential [21]. Its overexpression activates the HER2, PAK1, ERK, and Akt signaling pathways while suppressing p53 expression [23], creating a typical pro-proliferation and anti-apoptotic signaling environment. miR-145 inhibits MUC13 expression by directly targeting its 3'UTR [24], and this regulatory axis gradually becomes imbalanced during the progression from precancerous lesions to PDAC. Cucurbitacin D can restore miR-145 expression and reduce MUC13 and RRM1/2 levels, thereby reversing gemcitabine resistance [25]. This finding directly links MUC13 to the mechanisms of chemotherapy resistance.
In gastric cancer, MUC13 is highly expressed in intestinal-type gastric cancer and is almost absent in normal gastric mucosa [26]. Its spatial expression pattern varies among subtypes, with intestinal-type often showing apical localization, while diffuse-type exhibits diffuse cytoplasmic staining [26]. The long non-coding RNA BBOX1-AS1 competitively binds to miR-361-3p, relieving the inhibition of MUC13 [27], and the reduction of miR-132-3p also leads to increased MUC13 expression, activating the HER2, ERK, and Akt pathways [28]. In esophageal cancer, MUC13 enhances invasive ability by regulating O-glycan processing-related molecules [29]. Proteomic studies have shown its significant differential expression in gastric cancer patients with type 2 diabetes [30], suggesting its potential involvement in tumor progression associated with metabolic abnormalities.
In non-digestive system tumors, the cancer-promoting role of MUC13 is also evident. In lung cancer, it promotes cell proliferation and migration by activating ERK, JNK, and p38 signaling pathways [31]. In ovarian cancer, MUC13 synergizes with HER2 and PAK1 to enhance cell motility, with the JNK pathway involved in the formation of its tumorigenic phenotype [32]. In clear cell renal cell carcinoma (ccRCC), high expression of MUC13 is closely associated with poor prognosis [33,34]. It upregulates Cyclin D1, BCL-xL, and survivin through the NF-κB signaling pathway and reduces sensitivity to sorafenib and sunitinib [34]. Additionally, significant variant polymorphisms of MUC13 have been observed in the population with acute lymphoblastic leukemia [35], suggesting its potential genetic susceptibility significance. Overall, despite tissue-specific microenvironmental differences among various tumor types, MUC13 exhibits consistent molecular characteristics across multiple cancer types: upregulation of expression, loss of polarity, activation of signaling axes, and enhancement of drug resistance-related pathways. This cross-system consistency strengthens the theoretical foundation for its role as a broad-spectrum tumor biomarker and therapeutic target.
Currently, MUC13-targeted drugs are still in the preclinical stage, primarily focusing on gastrointestinal tumors such as pancreatic cancer and colorectal cancer. Various drug types (e. g., conjugated drugs, ADCs, small molecules, antibodies), led by American universities, are being developed with the mechanism of inhibiting MUC13, demonstrating the potential of this target in cancer therapy. Some of the pipelines under investigation are listed in the table below:
| Drug | Mechanism of action | Drug Type | Indications Under Research | Research Institutions | Highest R&D Stage |
|---|---|---|---|---|---|
| MUC13-SPIONS | MUC13 inhibitor | Conjugated drugs | Pancreatic cancer | University of Connecticut | Preclinical |
| TB-ADC-01 | MUC13 inhibitor | ADC | Rectal cancer | Theragen Bio Co., Ltd. | Preclinical |
| MUC13-SPION-CUR | MUC13 inhibitor | ADC | Pancreatic Ductal Adenocarcinoma | The University of Texas Rio Grande Valley | Preclinical |
| Cucurbitacin D | MUC13 inhibitor | Small molecule chemical drugs | Pancreatic Ductal Adenocarcinoma | The University of Tennessee Health Science Center | Preclinical |
| MUC13-targeted PPNPs (University of Texas Rio Grande Valley) | MUC13 inhibitor | Antibody | Pancreatic cancer | The University of Texas Rio Grande Valley | Preclinical |
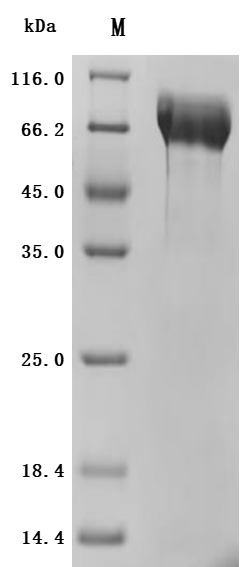
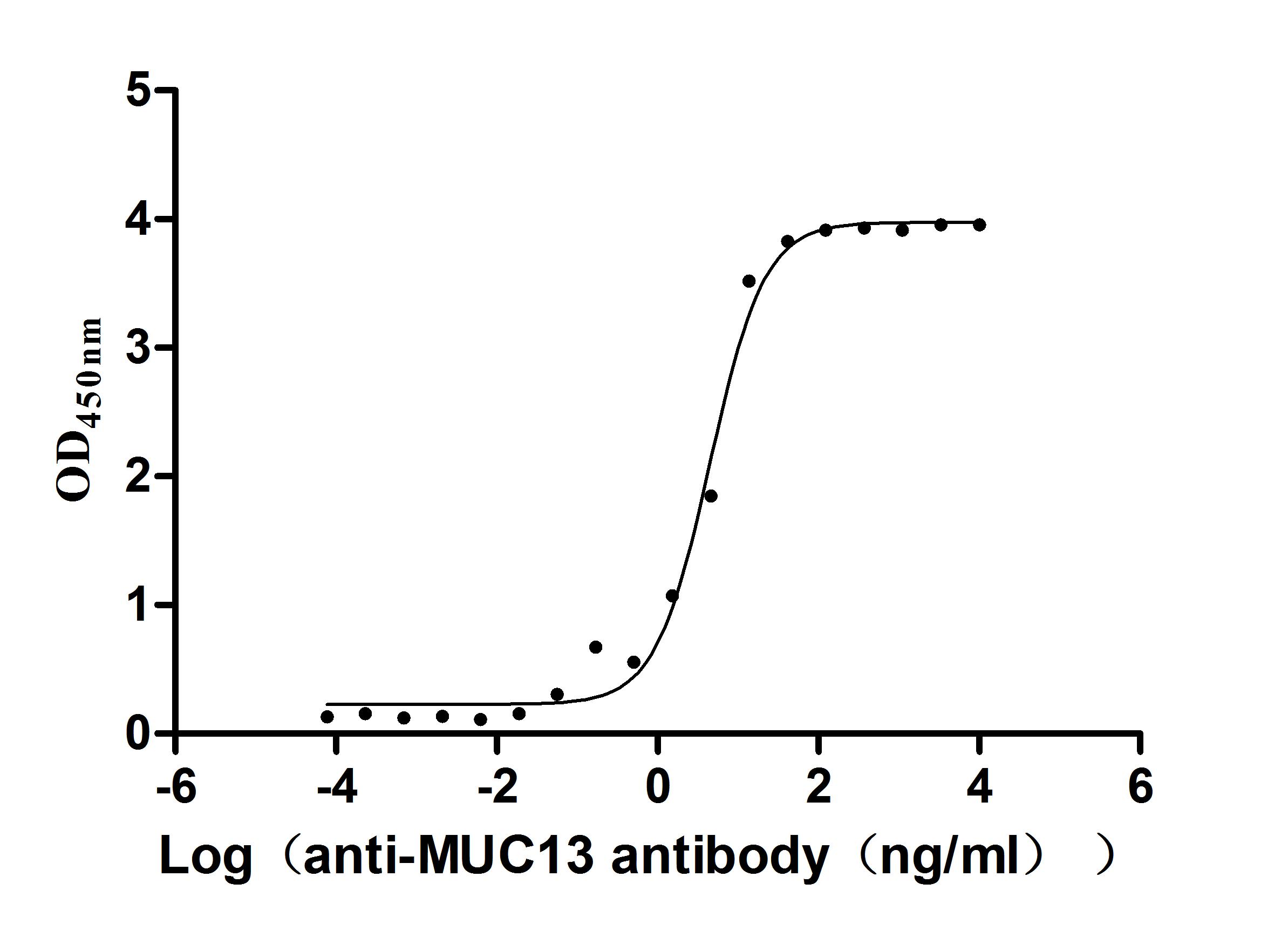
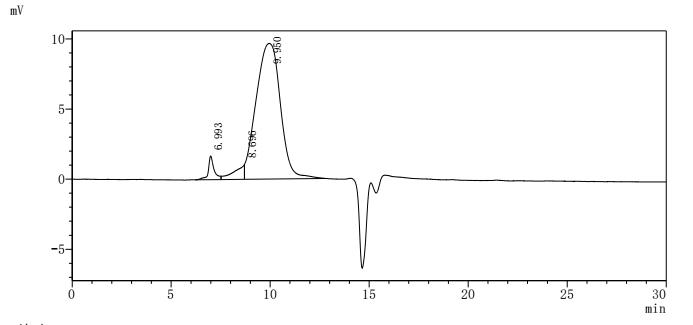
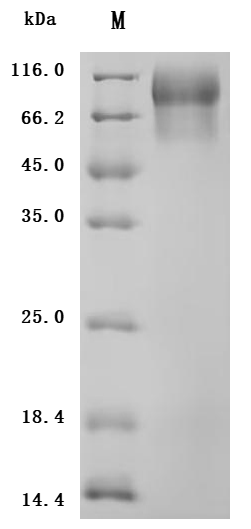
MUC13 has evolved from a traditional transmembrane mucin into a multi-pathway integrated oncogenic regulatory molecule. Its consistent pathogenic mechanism across multiple systemic malignancies positions it as a critical node linking epithelial barrier dysfunction with tumor signaling reprogramming. This also provides an important direction for diagnostic and therapeutic innovations in the context of precision medicine.Huamei Biology providesMUC13Recombinant proteins and antibody products, assisting you in related mechanism research and targeted drug development.
References
[1] Stephanie J. Williams, Daniel H. Wreschner, Mai H. Tran, Helen J. Eyre, Grant R. Sutherland, Michael A. McGuckin.(2001). MUC13, a Novel Human Cell Surface Mucin Expressed by Epithelial and Hemopoietic Cells.
[2] Nicolas Moniaux.(2001). Structural organization and classification of the human mucin genes.
[3] Peter Anthony.(2001). Mucins in the gastrointestinal tract in health and disease.
[4] Diane M. Maher, Brij K. Gupta, Satoshi Nagata, Meena Jaggi, Subhash C. Chauhan.(2011). Mucin 13: Structure, Function, and Potential Roles in Cancer Pathogenesis.
[5] Ryan J. King, Fang Yu, Pankaj K. Singh.(2017). Genomic alterations in mucins across cancers.
[6] Ranran Wang, Chao Yu, Dezhang Zhao, Mingjun Wu, Zhu Yang.(2013). The mucin-type glycosylating enzyme polypeptide N-acetylgalactosaminyltransferase 14 promotes the migration of ovarian cancer by modifying mucin 13.
[7] Sonam Kumari, Sheema Khan, Subash C. Gupta, Vivek K. Kashyap, Murali M. Yallapu, Subhash C. Chauhan, Meena Jaggi.(2017). Abstract 4399: MUC13 induced NFκB activation regulates metabolic reprograming by promoting its crosstalk with GLUT-1 receptor.
[8] Yong Sheng, Yaowu He, Sumaira Z. Hasnain, Ran Wang, Hui Tong, Daniel Clarke, Rohan Lourie, Iulia Oancea, K F Wong, John W. Lumley, Timothy H. Florin, Philip Sutton, John D. Hooper, Nigel A.J. McMillan, Michael A. McGuckin.(2016). Abstract 3564: MUC13 protects colorectal cancer cells from death by activating the NF-κb pathway and is a potential therapeutic target.
[9] Celia Segui‐Perez, Daphne A. C. Stapels, Ziliang Ma, Jinyi Su, Elsemieke M. Passchier, Bart Westendorp, Richard Wubbolts, Wei Wu, Jos P. M. van Putten, Karin Strijbis.(2024). MUC13 negatively regulates tight junction proteins and intestinal epithelial barrier integrity via protein kinase C.
[10] Xinyi Chen, Jingyao Tu, Mu Yang, Yuan Wang, Bo Liu, Hong Qiu, Xianglin Yuan.(2024). RUNX1-MUC13 Interaction Activates Wnt/β-Catenin Signaling Implications for Colorectal Cancer Metastasis.
[11] Kyle Doxtater, Manish K. Tripathi, Radhika Sekhri, Bilal Bin Hafeez, Sheema Khan, Nadeem Zafar, Stephen W. Behrman, Murali M. Yallapu, Meena Jaggi, Subhash C. Chauhan.(2023). MUC13 drives cancer aggressiveness and metastasis through the YAP1-dependent pathway.
[12] Shabnam Malik, Mohammed Sikander, Murali M. Yallapu, Subhash C. Chauhan.(2026). MUC13 is a crucial player in YAP1-β-catenin survival complex mediated cancer metastasis.
[13] Kyle Doxtater, Radhika Sekhri, Utkarsh K. Mishra, Meena Jaggi, Manish K. Tripathi, Subhash C. Chauhan.(2020). Abstract 4916: MUC13 enhances anchorage independent survival and cooperates with YAP1 and β-catenin towards colorectal cancer metastasis.
[14] Brij K. Gupta, Diane M. Maher, Mara C. Ebeling, Vasudha Sundram, Michael Koch, Douglas Lynch, Teresa Bohlmeyer, Akira Watanabe, Hiroyuki Aburatani, Susan E. Puumala, Meena Jaggi, Subhash C. Chauhan.(2012). Increased Expression and Aberrant Localization of Mucin 13 in Metastatic Colon Cancer.
[15] Brij K. Gupta, Diane M. Maher, Mara C. Ebeling, Rashmi Verma, Douglas Lynch, Michael Koch, Akira Watanabe, Hiroyuki Aburatani, Meena Jaggi, Subhash C. Chauhan.(2011). Abstract 1481: MUC13 expression enhances colon cancer progression.
[16] Brij K. Gupta, Diane M. Maher, Mara C. Ebeling, Douglas Lynch, Micheal D. Koch, Susan E. Puumala, Hiroyuki Aburatani, Meena Jaggi, Subhash C. Chauhan.(2012). Abstract 5325: Role of MUC13 in colon cancer progression.
[17] Kyle Doxtater, Kyle Doxtater.(2021). MUC13 Enhances Colorectal Cancer Metastasis.
[18] Ladislav Sojka, Alena Opattová, Linda Bartu, Josef Horak, Vlasta Korenková, Vendula Novosadová, Věra Křížková, Jan Brůha, Václav Liška, Michaela Schneiderová, Ondřej Kubeček, Ľudmila Vodičková, Markéta Urbanová, Jaromír Šimša, Pavel Vodička, Veronika Vymetálková.(2022).MUC13‑miRNA‑4647 axis in colorectal cancer: Prospects to identifications of risk factors and clinical outcomes.
[19] Bin Chen, Linguangjin Wu, Xiaoxia Tang, Ting Wang, Shuyun Wang, Hongjie Yu, Guangsheng Wan, Manli Xie, Ruijuan Zhang, Haijuan Xiao, Wanli Deng.(2022). RETRACTED: Quercetin Inhibits Tumorigenesis of Colorectal Cancer Through Downregulation of hsa_circ_0006990.
[20] Suresh Mathivanan, Justin Lim, Bow J. Tauro, Hong Ji, Robert L. Moritz, Richard J. Simpson.(2009). Proteomics Analysis of A33 Immunoaffinity-purified Exosomes Released from the Human Colon Tumor Cell Line LIM1215 Reveals a Tissue-specific Protein Signature.
[21] Anupam Dhasmana, Swati Dhasmana, Shivangi Agarwal, Sheema Khan, Shafiul Haque, Meena Jaggi, Murali M. Yallapu, Subhash C. Chauhan.(2023). Integrative big transcriptomics data analysis implicates crucial role of MUC13 in pancreatic cancer.
[22] Timon Vandamme, Marc Peeters, Fadime Dogan, Patrick Pauwels, Elvire Van Assche, Matthias Beyens, Geert Mortier, Geert Vandeweyer, Wouter W. de Herder, Guy Van Camp, Leo J. Hofland, Ken Op de Beeck.(2015). Whole-exome characterization of pancreatic neuroendocrine tumor cell lines BON-1 and QGP-1.
[23] Subhash C. Chauhan, Mara C. Ebeling, Diane M. Maher, Michael Koch, Akira Watanabe, Hiroyuki Aburatani, Yuhlong Lio, Meena Jaggi.(2011). MUC13 Mucin Augments Pancreatic Tumorigenesis.
[24] Sheema Khan, Mara C. Ebeling, Mohd Saif Zaman, Mohammed Sikander, Murali M. Yallapu, Neeraj Chauhan, Ashley M. Yacoubian, Stephen W. Behrman, Nadeem Zafar, Deepak Kumar, Paul A. Thompson, Meena Jaggi, Subhash C. Chauhan.(2014). MicroRNA-145 targets MUC13 and suppresses growth and invasion of pancreatic cancer.
[25] Mohammed Sikander, Shabnam Malik, Sheema Khan, Sonam Kumari, Neeraj Chauhan, Parvez Κhan, Fathi T. Halaweish, Bhavin Chauhan, Murali M. Yallapu, Meena Jaggi, Subhash C. Chauhan.(2019). Novel Mechanistic Insight into the Anticancer Activity of Cucurbitacin D against Pancreatic Cancer (Cuc D Attenuates Pancreatic Cancer).
[26] Takahiro Shimamura, Hirotaka Ito, Junji Shibahara, Akira Watanabe, Yoshitaka Hippo, Hirokazu Taniguchi, Yongxin Chen, Takeshi Kashima, Toshihiko Ohtomo, Fumihiko Tanioka, Hiroko Iwanari, Tatsuhiko Kodama, Teruhisa Kazui, Haruhiko Sugimura, Masashi Fukayama, Hiroyuki Aburatani.(2005). Overexpression of MUC13 is associated with intestinal‐type gastric cancer.
[27] Tao Cai, Binyu Peng, Jun Hu, Yan He.(2022). Long noncoding RNA BBOX1-AS1 promotes the progression of gastric cancer by regulating the miR-361-3p/Mucin 13 signaling axis.
[28] Liang He, Linlin Qu, Lijing Wei, Yan Chen, Jian Suo.(2017). Reduction of miR-132-3p contributes to gastric cancer proliferation by targeting MUC13.
[29] Guangxia Chen, Yi Han, Shiyu Liu, Guangqing Zhou, Xinxin Xu, Haihan Zhang, Zhentao Li, Chuannan Wu, Yulan Liu, Kai Fang, Gang Chen.(2021). MUC13 promotes the development of esophageal cancer by upregulating the expression of O-glycan process-related molecules.
[30] Hugo Osório, Cátia Silva, Marta Ferreira, Irene Gullo, Valdemar Máximo, Rita Barros, Fernando Mendonça, Carla Oliveíra, Fátima Carneiro.(2021). Proteomics Analysis of Gastric Cancer Patients with Diabetes Mellitus.
[31] Pang Yao, Yu Zhang, Hongyi Zhang, Wenhao Wang, Gang Jin, Jiawei Liu, Zijiang Zhu.(2021). MUC13 promotes lung cancer development and progression by activating ERK signaling.
[32] Subhash C. Chauhan, Kelley Vannatta, Mara C. Ebeling, Namita Vinayek, Akira Watanabe, Krishan K. Pandey, Maria C. Bell, Michael Koch, Hiroyuki Aburatani, Yuhlong Lio, Meena Jaggi.(2009). Expression and Functions of Transmembrane Mucin MUC13 in Ovarian Cancer.
[33] Zhiying Xu, Yidong Liu, Yuanfeng Yang, Jieti Wang, Guodong Zhang, Zheng Liu, Hangcheng Fu, Zewei Wang, Haiou Liu, Jiejie Xu.(2016). High expression of Mucin13 associates with grimmer postoperative prognosis of patients with non-metastatic clear-cell renal cell carcinoma.
[34] Yonghua Sheng, Choa Ping Ng, Rohan Lourie, Esha T. Shah, Yaowu He, Kuan Yau Wong, Inge Seim, Iulia Oancea, Christudas Morais, Penny L. Jeffery, John D. Hooper, Glenda C. Gobé, Michael A. McGuckin.(2017). MUC13 overexpression in renal cell carcinoma plays a central role in tumor progression and drug resistance.
[35] Angélica Leite de Alcântara, Lucas Favacho Pastana, Laura Patrícia Albarello Gellen, Giovana Miranda Vieira, Elizabeth Ayres Fragoso Dobbin, Thays Amâncio Silva, Esdras Edgar Batista Pereira, Williams Fernandes Barra, Juliana Carla Gomes Rodrigues, Marianne Rodrigues Fernades, Paulo Pimentel de Assumpção, João Farias Guerreiro, Sidney Emanuel Batista dos Santos, Ney Pereira Carneiro dos Santos.(2022). MUC family influence on acute lymphoblastic leukemia in Native American populations from Brazilian Amazon.


Comments
Leave a Comment