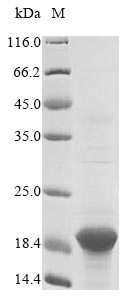
-B-SDS.jpg)
Call us
301-363-4651 (Available 9 a.m. to 5 p.m. CST from Monday to Friday)
β2-microglobulin (B2M), as the light chain subunit of MHC-I molecules, is a core component of the endogenous antigen presentation pathway and plays an irreplaceable role in CD8+ T cell recognition of viral and tumor antigens. In addition to classical immune functions, B2M is also involved in non-immune physiological processes such as neurocognitive regulation and metabolic homeostasis maintenance, and its abnormal expression is closely related to tumor immune escape, autoimmune diseases, neurodegenerative diseases, and cardiovascular risk. In recent years, the translational value of B2M as a predictive biomarker for immunotherapy response, an engineering target for universal cell therapy, and a new target for autoimmune disease intervention has become increasingly prominent. This review systematically summarizes the structural function, regulatory mechanisms, disease associations, and targeted therapy progress of B2M, providing a reference framework for related research and clinical translation.
2. Non-Immune Effects of Soluble B2M and Biomarker Value: Advantages and Limitations
3. B2M-MHC-I Antigen Presentation Mechanism and Regulation: From Assembly to Immune Escape
4. B2M-Related Signaling Pathways: Immune Cell Interaction and Tumor Endogenous Axis
5. B2M and Diseases: Immune Escape, Microenvironment, Monitoring, and Non-Tumor Pathology
B2M was first defined as the light chain of MHC-I molecules. Its classical function is to non-covalently bind to the MHC-I heavy chain (HC) to form a stable complex, maintain the structure of the peptide groove, and promote the assembly and presentation of endogenous peptides, thereby supporting CD8+ T cell recognition of viral and tumor antigens. Related work also emphasizes the importance of key pocket sites in the MHC domain and both ends of the peptide from the perspective of peptide presentation prediction, indirectly supporting the inherent association between "structural stability and presentation fate" [1]. At the evolutionary and mutational levels, B2M belongs to the basic components of IgSF, and its sequence conservation and functional optimization can be explained by topological and kinetic models; meanwhile, aggregation-susceptible mutations such as D76N have also been identified, which are consistent with amyloid deposition phenomena, suggesting that B2M has both a structural scaffold and potential pathological aggregation tendency [2].
Notably, recent evidence suggests that MHC-I heavy chains can appear on the cell surface in the form of B2M-free HC without binding to B2M and/or peptides, and further form homo/heterodimers; these molecules are upregulated after inflammatory stimulation or cell activation, have different glycosylation patterns from classical complexes, and may interact with KIR receptors, growth-promoting factors, and cytokines; in animal models, HLA-B27 induces spontaneous arthropathy in the context of B2M deficiency, suggesting that it may affect immune homeostasis and autoimmune phenotypes [3].
Overall, there is still a lack of systematic biochemical and structural evidence for the ontogeny, in vivo function of B2M-free HC, and its causal relationship with the efficiency of the classical MHC-I pathway; meanwhile, many computational and structural studies often do not incorporate the dynamic participation of B2M, glycosylation differences, and redox sensitivity into a unified framework, suggesting that future integration of structural and functional immunological evidence is needed to clarify its non-classical immunomodulatory potential [1,2,3].
Beyond immune function, the role of blood-borne B2M in the central nervous system provides a relatively continuous "association-intervention-mechanism" evidence chain: studies have linked elevated B2M in Down syndrome (DS) plasma to synaptic defects and memory impairment through paradigms such as parabiotic animals, plasma infusion, and systemic administration, and alleviated the phenotype by B2m gene deletion or anti-B2M antibody neutralization; it was further pointed out that B2M can bind to the S2 loop of the NMDAR subunit GluN1 and antagonize NMDAR function, and blocking with competitive short peptides can restore NMDAR-dependent synaptic function, thereby expanding B2M as an endogenous NMDAR antagonist and suggesting its direct pathogenic potential in cognitive impairment [4]. At the same time, extrapolation should be cautious: the comparability of systemic administration concentrations to human disease levels, the kinetic pathway of B2M affecting brain receptors, and the structural/affinity evidence of binding sites are still limited, which restrict direct translational inference [4].
From a biomarker perspective, B2M has cross-disease predictive potential, but there are challenges in specificity and causal interpretation. Large cohort data from the general population and non-CKD populations show a J-shaped non-linear correlation between B2M and all-cause mortality and cardiovascular death risk, suggesting that it may integrate multidimensional information such as chronic inflammation, immune activation, and renal clearance, and has certain risk stratification value [5]. However, observational studies are difficult to exclude residual confounding, and threshold determination and background correction are key limitations for clinical application [5]. In the CKD scenario, an antibody-free MRM mass spectrometry platform includes B2M in a multi-marker panel for absolute quantification, showing good correlation with ELISA, significant correlation with eGFR, and improved predictive ability for adverse outcomes in combined analysis, suggesting that standardized, multiplexed quantification strategies can partially overcome the limitations of single indicators [6]. Meanwhile, since B2M is strongly affected by renal clearance, its interpretation in non-nephropathy contexts needs to strictly distinguish between "elevation caused by decreased renal function" and "inflammatory/immune activation signals" [6].
Mature MHC-I consists of a heavy chain, B2M, and a bound peptide. B2M stabilizes heavy chain folding through non-covalent interactions and promotes the transport of the complex to the cell surface; B2M deletion/downregulation leads to loss of surface MHC-I expression and impaired CD8+ T cell recognition, forming the direct basis for immune escape [7,8]. The antigen processing and presentation machinery (APM) also relies on proteasome subunits (such as PSMB8/9/10) for peptide production, TAP1/2 transport, and ERAP modification, etc. Synergistic defects in multiple components significantly reduce the formation of nascent peptide-MHC-I complexes and are associated with immunotherapy resistance [8,9]. Functional studies further support causality: biallelic loss or CRISPR knockout of B2m can produce a resistant phenotype to PD-1 blockade in vivo, suggesting that APM disruption is sufficient to drive ICI failure in specific contexts [7].
At the regulatory level, inflammatory signals can enhance APM transcription: Type I interferon upregulates MHC-I, TAP1, PSMB8, and B2M through the STAT1/STAT2-IRF7 axis and transcriptional co-activators (including NLRC5), promoting antigen presentation and T cell activation [10,11]. Conversely, TGF-β downregulates multiple APM genes through the ALK5-Smad3 pathway, with temporal differences (e.g., ERAP1 decreases earlier, B2M decreases later), suggesting that a chronic immunosuppressive microenvironment may gradually form through this mechanism [9]. In addition, EZH2 can introduce H3K27me3 at HLA genes and B2M, TAP1 promoters to achieve sustained silencing, and suggests that EZH2 inhibition may restore APM expression and enhance immune clearance potential [12]. However, it should be emphasized that cross-tumor type heterogeneity and alternative immune pathways will modify the simple inference of "B2M deletion = inevitable ineffectiveness": for example, in MMRd tumors, although biallelic inactivation of B2M is common, ICI may still obtain anti-tumor activity through CD4+ T cell effects [13]; in addition, tumors with B2M or HLA mutations are often accompanied by higher potential neoantigen loads, but whether they can be converted into effective immune recognition through non-classical pathways still lacks sufficient functional evidence [14].
In the tumor microenvironment, B2M status and multiple signaling axes jointly shape the immunosuppressive landscape. Studies have shown that tumor-associated NK cells can drive enhanced myeloid-derived suppressor cell (MDSC) phenotypes, which are associated with IL-6 signaling and MDSC markers (such as S100A8/9, ARG1); intervention of the IL-6/STAT3 axis in animal models can alleviate T cell suppression and inhibit tumor growth, concentrating the key node of NK-MDSC interaction in the IL-6/STAT3 pathway [15]. Meanwhile, there is still a lack of quantitative evidence for the respective contributions of "contact-dependent receptors" and "secreted factors", which limits the predictability of generalization across tumor types [15].
B2M deletion may also have a direct impact on NK cell intrinsic signaling: knockout of B2M in the NK-92MI cell line results in decreased activation receptor expression, cytokine production, and cytotoxicity, accompanied by upregulated SHP-1 expression and phosphorylation, thereby inhibiting STAT3 and ERK phosphorylation; knockout of PTPN6 (SHP-1) can partially restore cytotoxicity, and CAR engineering can further overcome hyporesponsiveness, suggesting that SHP-1-mediated dephosphorylation is one of the inhibitory nodes [16]. However, this conclusion is mainly based on cell lines, and extrapolation to primary NK licensing mechanisms should be cautious, and SHP-1 is not the only negative regulatory factor [16].
In addition to immune interactions, B2M can also participate in tumor cell endogenous signaling: in glioblastoma, the interaction between B2M and PIP5K1A activates PI3K/AKT/mTOR and promotes MYC-mediated TGFβ1 secretion, thereby inducing M2-like polarization of macrophages, supporting tumor stemness and immunosuppressive microenvironment [17]. Therefore, the generalization of B2M function needs to explicitly incorporate tissue and cell lineage differences [17].
Large-scale colorectal cancer genomic studies have shown that MSI-high tumors often exhibit immune editing features of the antigen presentation machinery, including biallelic loss of B2M and HLA genes (including copy number alterations and copy-neutral LOH) [18]. Clinical cohorts further report that the B2M mutation rate in MSI-H CRC can reach approximately 24%, and mutations are highly consistent with protein expression loss, supporting the close association between "genotype and phenotype" [19]; meanwhile, B2M hotspot mutations are enriched in microsatellite unstable CRC, and may exist before immunotherapy, suggesting that they may be both early evolutionary events and selected under therapeutic pressure [20].
In other tumors, B2M alterations co-occur with phenotypes such as reduced HLA expression and decreased CD8+ T cell infiltration, reinforcing the explanatory framework of "impaired antigen presentation → immune escape", for example, B2M mutations in EBV-positive DLBCL coexist with reduced HLA expression and decreased CD8+ infiltration [21], and are consistent with the pattern of frequent impairment of HLA-related genes in multiple EBV-related or hematological tumors [22]. In malignant gliomas, studies on the complexity of HLA region sequencing have also identified somatic mutations in B2M and TAP genes, and suggest that they may be enriched in recurrent GBM, pointing to the role of selective pressure during progression or treatment [23]. In MSI/dMMR pancreatic ductal adenocarcinoma, B2M inactivation can exhibit heterogeneity between primary and metastatic lesions, suggesting that treatment evaluation needs to focus on spatial differences and the importance of metastatic sites [24].
IIn DLBCL, mutations in genes such as B2M are associated with multiple extranodal involvement (ENI), worse prognosis, enhanced Treg recruitment at the TME level, and reduced stromal gene expression; animal models also suggest that B2M mutations are associated with bone marrow invasion, implying that they may drive invasive phenotypes through decreased antigen presentation and altered immune infiltration profiles [25]. However, the current understanding of how they directly drive Treg recruitment (such as chemotactic networks or stromal remodeling) remains largely correlative.
In Lynch syndrome-related CRC, even in the presence of tumor cell B2M loss, PD-L1 upregulation and abundant CD3+/CD8+ infiltration can still be observed, suggesting that B2M deletion does not necessarily equate to "complete immune evasion", and ICI sensitivity should be interpreted in combination with a broader immune landscape [26]. In addition, integrated genomic studies of PMBL show that multiple immune escape-related genes including B2M are frequently mutated and coexist with JAK-STAT and NF-κB alterations, supporting immune evasion as one of the core mechanisms of this subtype [27]; single-cell studies of follicular lymphoma have also identified a gene set associated with the immune checkpoint co-expression network in Treg cells, including B2M, suggesting that it may have significance in immune cell endogenous expression and regulation, but direct functional verification is still lacking [28].
Whole-genome sequencing of sorted HRS cells from cHL reveals complex structural variations and evolutionary timelines: mutations in immune escape-related genes including B2M often occur before large-scale chromosomal gains, supporting them as early key events from a temporal perspective [29]. At the more accessible clinical monitoring level, rapid gene panel studies of ctDNA show that the overall variant detection rate at diagnosis is relatively high, and the B2M mutation rate is reported to be approximately 33.3%; ctDNA rapidly turns negative early in treatment and is highly consistent with PET indicators, suggesting that it can be used as a dynamic biomarker complementary to imaging [30]. However, "undetectable" is limited by the limit of detection, tumor DNA release, and panel coverage, and MRD and clonal evolution monitoring may still have blind spots.
In addition, B2M also appears in DLBCL genetic classification (e.g., one of the common variants in the EZB-like subtype), suggesting that it has certain universality across lymphoma subtypes and provides potential clues for classification and immune-related decision-making [31].
In pSS salivary gland microarray differential expression and network analysis, B2M is significantly upregulated and ranks among the hub genes, with upregulated genes enriched in antigen presentation and interferon pathways, suggesting that it is closely related to local immune activation, but functional verification is still needed to distinguish between driving and accompanying markers [32]. Single-cell studies of RA peripheral blood have observed downregulation of multiple genes including B2M in the DNT cell population, accompanied by suppressed oxidative phosphorylation and impaired antigen presentation, suggesting that decreased B2M in certain immune cell subsets may parallel decreased metabolic and presentation functions, but the consistency between transcription and protein/surface expression still needs experimental confirmation [33]. Proteomic studies of KD gingival crevicular fluid have identified upregulated proteins such as B2M and suggested their potential as early diagnostic biomarkers, but the causal pathway between local fluid sources and systemic pathology remains to be clarified [34]. In addition, dialysis can significantly reduce circulating B2M levels, suggesting that in populations with impaired renal function or receiving dialysis intervention, B2M changes may be strongly affected by treatment operations, constituting an important confounding factor [35].
In recent years, the research and development of drugs targeting β2-microglobulin (B2M) has shown a diversified trend, with indications expanding from traditional tumor immunotherapy to autoimmune diseases and neurodegenerative diseases. Currently, the fastest progressing projects focus on fields such as melanoma and diabetes, while innovative therapies based on gene editing and universal CAR-T technology have also entered the clinical verification stage, demonstrating the translational potential of B2M as a multifunctional target. Some of the research pipelines are summarized in the table below:
| Drug Name | Type | Indications | Development Stage | Research Institution |
|---|---|---|---|---|
| B2M monoclonal antibody | Monoclonal antibody | Melanoma | Preclinical | Multiple research institutions |
| B2M inhibitor | Small molecule compound | Type 1 diabetes | Preclinical | Multiple institutions |
| B2M gene-edited CAR-T | Cell therapy | Hematological malignancies | Phase 1 | Multiple clinical centers |
| Universal CAR-T (B2M knockout) | Cell therapy | Solid tumors/hematological malignancies | Phase 1/2 | Biotechnology companies |
| B2M neutralizing antibody | Antibody drug | Alzheimer's disease | Exploratory stage | Research institutions |
| B2M vaccine | Vaccine | Tumor immunotherapy | Preclinical | Pharmaceutical company collaboration projects |
B2M occupies a key node in the "antigen presentation - immune escape - microenvironment remodeling - clinical monitoring - therapeutic engineering" chain: its deletion or mutation can impair CD8+ T cell recognition through impaired MHC-I assembly, and is associated with immunotherapy resistance in multiple tumor types. CUSABIO provides B2M recombinant proteins, antibodies, and ELISA kits to support your mechanistic research and targeted drug development.
● B2M Recombinant Proteins
● B2M Antibodies
● B2M ELISA Kits
References
[1] Hans-Christof Gasser, Georges Bedran, Bo Ren, David Goodlett, Javier Alfaro, Ajitha Rajan.(2021). Interpreting BERT architecture predictions for peptide presentation by MHC class I proteins.
[2] J. C. Phillips.(2022). Evolution and Mutations of Beta 2 Microglobulin.
[3] Mepur H Ravindranath, Narendranath M Ravindranath, Senthamil R Selvan, Fatiha El Hilali, Carly J Amato-Menker, Edward J Filippone.(2023). Cell Surface B2m-Free Human Leukocyte Antigen (HLA) Monomers and Dimers: Are They Neo-HLA Class and Proto-HLA?.
[4] Yue Gao, Yujuan Hong, Lihong Huang, Shuang Zheng, Haibin Zhang, Shihua Wang, Yi Yao, Yini Zhao, Lin Zhu, Qiang Xu, Xuhui Chai, Yuanyuan Zeng, Yuzhe Zeng, Liangkai Zheng, Yulin Zhou, Hong Luo, Xian Zhang, Hongfeng Zhang, Ying Zhou, Guo Fu, Hao Sun, Timothy Y Huang, Qiuyang Zheng, Huaxi Xu, Xin Wang.(2023). β2-microglobulin functions as an endogenous NMDAR antagonist to impair synaptic function..
[5] Hang Fang, Qiankun Zhang, Lie Jin.(2023). Association of beta-2-microglobulin with cardiovascular and all-cause mortality in the general and non-CKD population..
[6] Manousos Makridakis, Georgia Kontostathi, Eleni Petra, Rafael Stroggilos, Vasiliki Lygirou, Szymon Filip, Flore Duranton, Harald Mischak, Angel Argiles, Jerome Zoidakis, Antonia Vlahou.(2020). Multiplexed MRM-based protein quantification of putative prognostic biomarkers for chronic kidney disease progression in plasma..
[7] Scott Gettinger, Jungmin Choi, Katherine Hastings, Anna Truini, Ila Datar, Ryan Sowell, Anna Wurtz, Weilai Dong, Guoping Cai, Mary Ann Melnick, Victor Y. Du, Joseph Schlessinger, Sarah B. Goldberg, Anne C. Chiang, Miguel F. Sanmamed, Ignacio Melero, Jackeline Agorreta, Luis M. Montuenga, Richard P. Lifton, Soldano Ferrone, Paula Kavathas, David L. Rimm, Susan M. Kaech, Kurt A. Schalper, Roy S. Herbst, Katerina Politi.(2017). Impaired HLA Class I Antigen Processing and Presentation as a Mechanism of Acquired Resistance to Immune Checkpoint Inhibitors in Lung Cancer.
[8] Karolina Žilionytė, Ugnė Bagdzevičiūtė, Agata Mlynska, Elena Urbštaitė, Emilija Paberalė, Neringa Dobrovolskienė, Jan Aleksander Krasko, Vita Pašukonienė.(2022). Functional antigen processing and presentation mechanism as a prerequisite factor of response to treatment with dendritic cell vaccines and anti-PD-1 in preclinical murine LLC1 and GL261 tumor models..
[9] Alix K. Berglund, Anna L. Hinson, Lauren V. Schnabel.(2023). TGF-β downregulates antigen processing and presentation genes and MHC I surface expression through a Smad3-dependent mechanism.
[10] Hemin Jiang, Yue Li, Min Shen, Yucheng Liang, Qian Yu, Hao Dai, Kuanfeng Xu, Xinyu Xu, Hui Lv, Jie Zhang, Tao Yang, Qi Fu.(2022). Interferon‐α promotes MHC I antigen presentation of islet β cells through STAT1‐IRF7 pathway in type 1 diabetes.
[11] Chrissie E B Ong, Amanda L Patchett, Jocelyn M Darby, Jinying Chen, Guei-Sheung Liu, A Bruce Lyons, Gregory M Woods, Andrew S Flies.(2021). NLRC5 regulates expression of MHC-I and provides a target for anti-tumor immunity in transmissible cancers..
[12] Wei Ding, Zihan Ding, Qinghong Zeng, Yan Qiu, Christopher R Donnelly, Yuqi Wu, Yuchen Jiang, Qi Han, Hao Xu, Hao Cui, Xiangfei Liu, Xin Chen, Sixin Jiang, Mei Huang, Dan Pan, Dan Yang, Li Li, Lihong Yao, Minghai Tang, Jing Li, Taiwen Li, Xiaoping Xu, Qianming Chen, Hang Zhao, Longyu Li, Lu Jiang, Xiaobo Luo.(2025). Prevention of cancer initiation by augmenting MHC-I antigen presentation via EZH2 inhibition..
[13] Giovanni Germano, Steve Lu, Giuseppe Rospo, Simona Lamba, Benoı̂t Rousseau, Sonia Fanelli, Denise Stenech, Dung T. Le, John L. Hays, Maria Grazia Totaro, Vito Amodio, Rosaria Chilà, Anna Mondino, Luis A. Díaz, Federica Di Nicolantonio, Alberto Bardelli.(2021). CD4 T Cell–Dependent Rejection of Beta-2 Microglobulin Null Mismatch Repair–Deficient Tumors.
[14] Andrea Castro, Kıvılcım Öztürk, Rachel Marty Pyke, Xian Su, Maurizio Zanetti, Hannah Carter.(2019). Elevated neoantigen levels in tumors with somatic mutations in the HLA-A, HLA-B, HLA-C and B2M genes.
[15] Shiyong Neo, Le Tong, Joni Chong, Yaxuan Liu, Jing Xu, Mariana M.S. Oliveira, Yi Chen, Ziqing Chen, Keene Lee, Nutsa Burduli, Xinsong Chen, Juan Gao, Ran Ma, Jia Pei Lim, Jianxin Huo, Shengli Xu, Evren Alici, Stina L. Wickström, Felix Haglund, Johan Hartman, Arnika Kathleen Wagner, Yihai Cao, Rolf Kiessling, Kong‐Peng Lam, Lisa S. Westerberg, Andreas Lundqvist.(2024). Tumor-associated NK cells drive MDSC-mediated tumor immune tolerance through the IL-6/STAT3 axis.
[16] Kuo Yu, Xiaolong Liu, Guangyuan Wu, Zhongyao An, Xin Wang, Yang Liu, Hailong Wang, Mingli Huang, Linlin Zhao, Ce Shi, Xin Sun, Lu Xu, Sen Qi, Xin Zhang, Yueqiu Teng, Song Guo Zheng, Zhiren Zhang, Zhenkun Wang.(2025). Targeting SHP-1-Mediated Inhibition of STAT3 and ERK Signalling Pathways Rescues the Hyporesponsiveness of MHC-I-Deficient NK-92MI..
[17] Daqi Li, Qian Zhang, Lu Li, Kexin Chen, Junlei Yang, Deobrat Dixit, Ryan C Gimple, Shusheng Ci, Chenfei Lu, Lang Hu, Jiancheng Gao, Danyang Shan, Yangqing Li, Junxia Zhang, Zhumei Shi, Danling Gu, Wei Yuan, Qiulian Wu, Kailin Yang, Linjie Zhao, Zhixin Qiu, Deguan Lv, Wei Gao, Hui Yang, Fan Lin, Qianghu Wang, Jianghong Man, Chaojun Li, Weiwei Tao, Sameer Agnihotri, Xu Qian, Yu Shi, Yongping You, Nu Zhang, Jeremy N Rich, Xiuxing Wang.(2022). β2-Microglobulin Maintains Glioblastoma Stem Cells and Induces M2-like Polarization of Tumor-Associated Macrophages..
[18] Catherine S. Grasso, Marios Giannakis, Daniel K. Wells, Tsuyoshi Hamada, Xinmeng Jasmine Mu, Michael J. Quist, Jonathan A. Nowak, Reiko Nishihara, Zhi Rong Qian, Kentaro Inamura, Teppei Morikawa, Katsuhiko Nosho, Gabriel Abril-Rodríguez, Charles M. Connolly, Helena Escuin-Ordinas, Milan S. Geybels, William M. Grady, Li Hsu, Siwen Hu‐Lieskovan, Jeroen R. Huyghe, Yeon Joo Kim, Paige Krystofinski, Mark D.M. Leiserson, Dennis Montoya, Brian B. Nadel, Matteo Pellegrini, Colin C. Pritchard, Cristina Puig-Saus, Elleanor H. Quist, Benjamin J. Raphael, Stephen J. Salipante, Daniel Sanghoon Shin, Eve Shinbrot, Brian H. Shirts, Sachet A. Shukla, Janet L. Stanford, Wei Sun, Jennifer Tsoi, Alexander Upfill‐Brown, David A. Wheeler, Catherine J. Wu, Ming Yu, Syed Hassan Ejaz Zaidi, Jesse M. Zaretsky, Stacey Gabriel, Eric S. Lander, Levi A. Garraway, Thomas J. Hudson, Charles S. Fuchs, Antoni Ribas, Shuji Ogino, Ulrike Peters.(2018). Genetic Mechanisms of Immune Evasion in Colorectal Cancer.
[19] Sumit Middha, Rona Yaeger, Jinru Shia, Zsofia K. Stadler, Sarah King, Shanna Guercio, Viktoriya Paroder, David D. B. Bates, Satshil Rana, Luis A. Díaz, Leonard B. Saltz, Neil H. Segal, Marc Ladanyi, Ahmet Zehir, Jaclyn F. Hechtman.(2019). Majority ofB2M-Mutant and -Deficient Colorectal Carcinomas Achieve Clinical Benefit From Immune Checkpoint Inhibitor Therapy and Are Microsatellite Instability-High.
[20] Su Yeon Yeon, Seung-Hyun Jung, Yun Sol Jo, Eun Ji Choi, Min Sung Kim, Yeun-Jun Chung, Sug Hyung Lee.(2019). Immune checkpoint blockade resistance-related B2M hotspot mutations in microsatellite-unstable colorectal carcinoma..
[21] Miaoxia He, Bin Liu, Gusheng Tang, Lijuan Jiao, Xuefei Liu, Shuyi Yin, Tao Wang, Jie Chen, Lei Gao, Xiong Ni, Libin Wang, Lili Xu, Jianmin Yang.(2022). B2M mutation paves the way for immune tolerance in pathogenesis of Epstein-Barr virus positive diffuse large B-cell lymphomas..
[22] Chantana Polprasert, Yasuhide Takeuchi, Hideki Makishima, Kitsada Wudhikarn, Nobuyuki Kakiuchi, Nichthida Tangnuntachai, Thamathorn Assanasen, Wimonmas Sitthi, Hamidah Muhamad, Panisinee Lawasut, Sunisa Kongkiatkamon, Udomsak Bunworasate, Koji Izutsu, Yuichi Shiraishi, Kenichi Chiba, Hiroko Tanaka, Satoru Miyano, Seishi Ogawa, Kenichi Yoshida, Ponlapat Rojnuckarin.(2020). Frequent mutations in HLA and related genes in extranodal NK/T cell lymphomas.
[23] Sara C Schulte, Wolfgang Peter, Georg Rosenberger, Moritz Schäfer, Cecile L Maire, Alessandra Rünger, Alice Ryba, Kristoffer Riecken, Krystian D Fita, Jakob Matschke, Nuray Akyüz, Judith Dierlamm, Gunnar W Klau, Franz L Ricklefs, Jens Gempt, Manfred Westphal, Katrin Lamszus, Alexander Dilthey, Malte Mohme.(2025). Somatic Mutations in HLA Class Genes and Antigen-Presenting Molecules in Malignant Glioma..
[24] Claudio Luchini, Andrea Mafficini, Deyali Chatterjee, Maria L Piredda, Concetta Sciammarella, Pooja Navale, Giuseppe Malleo, Paola Mattiolo, Giovanni Marchegiani, Antonio Pea, Roberto Salvia, Lodewijk A Brosens, Gaetano Paolino, Maria G Mastrosimini, Nicola Silvestris, Michele Milella, Liang Cheng, Volkan N Adsay, Rita T Lawlor, Aldo Scarpa.(2022). Histo-molecular characterization of pancreatic cancer with microsatellite instability: intra-tumor heterogeneity, B2M inactivation, and the importance of metastatic sites..
[25] Rong Shen, Pengpeng Xu, Nan Wang, Hongmei Yi, Lei Dong, Di Fu, Jinyan Huang, Hengye Huang, Anne Janin, Shu Cheng, Li Wang, Weili Zhao.(2020). Influence of oncogenic mutations and tumor microenvironment alterations on extranodal invasion in diffuse large B‐cell lymphoma.
[26] Joanna Walkowska, Thomas Kallemose, Göran Jönsson, Mats Jönsson, Ove Andersen, Mads Hald Andersen, Inge Marie Svane, Anne Langkilde, Mef Nilbert, Christina Therkildsen.(2018). Immunoprofiles of colorectal cancer from Lynch syndrome.
[27] Anja Mottok, Stacy Hung, Elizabeth A. Chavez, Bruce W. Woolcock, Adèle Telenius, Lauren C. Chong, Barbara Meissner, Hisae Nakamura, Christopher Rushton, Elena Viganò, Clémentine Sarkozy, Randy D. Gascoyne, Joseph M. Connors, Susana Ben‐Neriah, Andrew J. Mungall, Marco A. Marra, Reiner Siebert, David W. Scott, Kerry J. Savage, Christian Steidl.(2019). Integrative genomic analysis identifies key pathogenic mechanisms in primary mediastinal large B-cell lymphoma.
[28] Noemi Andor, Erin F. Simonds, Debra K. Czerwinski, Jiamin Chen, Susan M. Grimes, Christina Wood-Bouwens, Grace Zheng, Matthew Kubit, Stephanie Greer, William A. Weiss, Ronald Levy, Hanlee P. Ji.(2018). Single-cell RNA-Seq of follicular lymphoma reveals malignant B-cell types and coexpression of T-cell immune checkpoints.
[29] Francesco Maura, Bachisio Ziccheddu, Jenny Xiang, Bhavneet Bhinder, Joel Rosiene, Federico Abascal, Kylee Maclachlan, Kenneth Wha Eng, Manik Uppal, Feng He, Wei Zhang, Qi Gao, Venkata D. Yellapantula, Vicenta Trujillo-Alonso, Sunita I. Park, Matthew J. Oberley, Elizabeth Ruckdeschel, Megan S. Lim, Gerald Wertheim, Matthew J. Barth, Terzah M. Horton, Andriy Derkach, Alexandra E. Kovach, Christopher J. Forlenza, Yanming Zhang, Ola Landgren, Craig H. Moskowitz, Ethel Cesarman, Marcin Imieliński, Olivier Elemento, Mikhail Roshal, Lisa Giulino‐Roth.(2023). Molecular Evolution of Classic Hodgkin Lymphoma Revealed Through Whole-Genome Sequencing of Hodgkin and Reed Sternberg Cells.
[30] Vincent Camus, Mathieu Viennot, Justine Lequesne, Pierre‐Julien Viailly, Élodie Bohers, Lucile Bessi, B. Marcq, Pascaline Etancelin, Sydney Dubois, Jean‐Michel Picquenot, Elena‐Liana Veresezan, Marie Cornic, Lucie Burel, Justine Loret, Stéphanie Becker, Pierre Decazes, Pascal Lenain, Stéphane Leprêtre, Emilie Lemasle, Hélène Lanic, Anne‐Lise Ménard, Nathalie Contentin, Hervé Tilly, Aspasia Stamatoullas, Fabrice Jardin.(2020). Targeted genotyping of circulating tumor DNA for classical Hodgkin lymphoma monitoring: a prospective study.
[31] Rong Shen, Di Fu, Lei Dong, Mu‐Chen Zhang, Qing Shi, Zi‐Yang Shi, Shu Cheng, Li Wang, Pengpeng Xu, Weili Zhao.(2023). Simplified algorithm for genetic subtyping in diffuse large B-cell lymphoma.
[32] Lei Zhang, Poshi Xu, Xiaoyu Wang, Zongshan Zhang, Wenxin Zhao, Zhengmin Li, Guangxia Yang, Panpan Liu.(2019). Identification of differentially expressed genes in primary Sjögren's syndrome.
[33] Xinqiang Song, Yu Zhang, Lijun Zhao, Jinke Fan, Tao Peng, Ying Ma, Nailiang Guo, Xiaotong Wang, Xudong Liu, Zhe Liu, Lei Wang.(2023). Analyzation of the Peripheral Blood Mononuclear Cells Atlas and Cell Communication of Rheumatoid Arthritis Patients Based on Single-Cell RNA-Seq.
[34] Xue Fan, Ying Li, Yuehao Xu, Jianqing Lin, Xin Guo, Jinwen Liao, Mingguo Xu.(2025). New biomarkers of Kawasaki disease identified by gingival crevicular fluid proteomics..
[35] Jan Kampmann, Michael Mejer Hunderup, Eva Rabing Brix Petersen, Vivi Andersen, Thor Aage Skovsted.(2024). High-sensitive troponin T, suPAR and Beta-2-microglobulin changes in concentration during hemodialysis.


Comments
Leave a Comment